
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
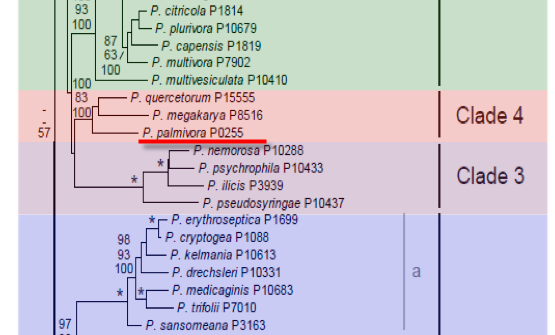
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora palmivora var. palmivora (E.J. Butler) E.J. Butler 1919[1918] (Oomycetes, Pythiales) Pythium palmivorum E.J. Butler 1907Phytophthora palmivora (E.J. Butler) E.J. Butler 1919 [1918]
Pythium palmivorum E.J. Butler 1907Phytophthora palmivora (E.J. Butler) E.J. Butler 1919 [1918]
= Kawakamia carica Hara 1915
Phytophthora carica (Hara) Hori 1916
= Phytophthora faberi Maubl. 1909
= Phytophthora fici Hori 1915 Note: Based on P. carica, see Waterhouse 1963. Phytophthora fici Venkata Rao 1916 is an invalid name, nom. nud. (Waterhouse 1963, p. 189.
= Phytophthora omnivora var. arecae L.C. Coleman 1910
Phytophthora arecae (L.C. Coleman) Pethybr. 1913
Phytophthora cactorum var. arecae (L.C. Coleman) Sacc. & Trotter 1912
= Phytophthora theobromae L.C. Coleman 1910
Phytophthora palmivora var. theobromae (L.C. Coleman) Orellana 1959
Notes: Phytophthora magakarya and Phytophthora capsici were previously referred to as P. palmivora, but are now considered distinct species (Erwin & Ribeiro 1996). Erwin & Ribeiro (1996) erroneously listed Phythophthora omnivora de Bary 1881 as a synonym, but it is instead a superfluous name for P. cactorum (see Waterhouse 1963). P. arecae was until recently considered to be a distinct species, but is now considered to be synonymous with P palmivora as originally suggested by Tucker (1931) (Mitchell & Kannwischer-Mitchell 1992, see ITS analysis in Cooke et al. 2000).
Distribution: Cosmopolitan.
Substrate: Leaves, buds, stems, fruits, roots.
Disease Note: Black pod and canker of cacao; patch canker, black stripe and leaf fall of rubber; bud rot of coconut, fruit and stem rot of pawpaw; root rot and damping-off of seedlings.
Host: Over 138 species from various plant families (Stamps 1985).
Supporting Literature:
Cooke, D.E.L., Drenth, A., Duncan, J.M., Wagels, G., and Brasier, C.M. 2000. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genet. Biol. 30: 17-32
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Stamps, D.J. 1985. Phytophthora palmivora. C.M.I. Descript. Pathog. Fungi Bact. 831: 1-2
Tucker, C.M. 1931. Taxonomy of the genus Phytophthora de Bary. Univ. Missouri Agric. Exp. Sta. Bull. 153: 1-208
Waterhouse, G.M. 1963. Key to the species of Phytophthora de Bary. Mycol. Pap. 92: 1-22
Updated on May 26, 2006
Since the morphological and physiological characteristics of P. palmivora vary considerably, precise descriptions of sporangia and oospores of all forms and strains cannot be made under one heading. A range in the variation is to be expected for isolates from different hosts (Table 2). See Brasier and Griffin (1979) for excellent photomicrographs of sporangia and oospores.
P. palmivora is classified in group II (Stamps et al. 1990) and described in detail by Waterhouse (1974b) and Stamps (1985a). The dimensions of sporangia, chlamydospores, sex organs, and growth temperatures for several isolates from different hosts are summarized in Table 2. Characteristics of P. palmivora, P. megakarya, and P. capsici are compared in Table 4. See Table 4.2 and Table 4.3 for tabular keys and Appendix 4.9 for a dichotomous key in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). Morphology is shown in Figure 1. Brasier and Griffin (1979) illustrate the wide variation in shapes of sporangia. Dimensions, shapes, and caducity of sporangia from several different reports on P. palmivora, P. megakarya, P. capsici, P. arecae, P. meadii, P. botryosa, P. heveae, and P. nicotianae (P. parasitica) are given in Table 1. Table 3 compares the chlamydospore and sex organ characteristics for these species.
1. Sporangia
Sporangia are variable in shape, depending on the isolate, mostly elliptical to ovoid, prominently papillate, and form on sympodial sporangiophores (Figure 1, 4.9C, 4.12E, and 4.16A in Phytophthora Diseases Worldwide [Erwin and Ribeiro 1996]). As many as 20 sporangia can occur on one sympodium. Sporangia are caducous (deciduous) and after they fall off the sporangiophore are characterized by short (5-µm) pedicels. Sporangia are variable in size (Table 2) but average 40 to 60 µm in length and 25 to 35 µm in breadth with a length-breadth ratio of 1.4 to 2.0 (Holliday 1980). Isolates from cocoa had a length-breadth ratio of 1.67 to 1.93 (Brasier and Griffin 1979). A recent study of a wide range of isolates from several hosts showed that sporangia were ellipsoid to spherical (predominantly ellipsoid with a broad base) with average lengths of 45.3 µm and breadths of 29.8 µm. The length-breadth ratio averaged 1.5 (range 1.3 to 1.8) (Mchau and Coffey 1994a; Table 1). About 10 to 40 zoospores are discharged from sporangia incubated in water (Holliday 1980). Sporangia are stimulated by light (Brasier 1969a). According to Waterhouse (1974b), sporangia are more elongated in light.
2. Chlamydospores
Globose to subglobose chlamydospores are terminal or intercalary on mycelium of most isolates of P. palmivora (Figure 4.24C and D); however, six of 93 isolates from widely distributed areas did not form chlamydospores on V8 juice agar (Mchau and Coffey 1994a). Chlamydospore diameters measure 32 to 42 µm (Holliday 1980), average 33 µm (Waterhouse 1974b), 36 µm (Ashby 1929a), and 36.2  9.6 µm (Mchau and Coffey 1994a) (Table 2).
9.6 µm (Mchau and Coffey 1994a) (Table 2).
3. Sex Organs
P. palmivora forms oogonia and oospores when A1 and A2 mating strains are paired (Ashby 1929; Brasier and Griffin 1979). Oogonia are spherical, and antheridia are amphigynous (Waterhouse 1974a). The mean oogonial diameter ranges from 22.3 to 34.8 µm (Brasier and Griffin 1979). Oospores average 22.8 0.1 µm (Dantanarayana et al. 1984) (Table 3). Oospores are loose in the oogonia (aplerotic). The distribution of A1 and A2 mating types is reported by Zentmyer et al. (1973) and Zentmyer (1974).
Babacauh (1983) made crosses of a P. palmivora A2 isolate with a \'near P. citrophthora\' A1 isolate, both from cocoa, and noted that amphigyny may result from the ability of the same hyphal tip to differentiate both an antheridium and an oogonium.
4. Distinguishing Characteristics
P. palmivora is distinguished from other heterothallic species with conspicuous papillate sporangia by production of chlamydospores and elongated, ellipsoidal to ovoid deciduous sporangia with short pedicels (Ho 1990). Table 3 and Table 4 summarize some of the distinguishing characteristics. Ho agrees with Waterhouse (1963) that P. carica, which has been reported on fig (Ficus carica L.) in Taiwan, is a synonym of P. palmivora. The review of Dantanarayana et al. (1984) on the taxonomy of Phytophthora species isolated from rubber in Sri Lanka indicates that P. meadii, the principal pathogen on rubber, is distinguished from P. palmivora by its nearly spherical sporangia compared with the ovoid sporangia of P. palmivora. Pedicels of P. meadii sporangia are medium in length (mean of 12.3 µm), while pedicels of P. palmivora are short (mean of 3.2 µm). Chlamydospores are rare on P. meadii but abundant on P. palmivora. Both species are heterothallic. The morphology of sporangia and patterns of cultures for these species are illustrated by Dantanarayana et al. (1984). Tucker (1931) considered P. arecae and P. meadii to be synonymous with P. palmivora. Oudemans and Coffey (1991b, c) found that P. arecae and P. palmivora are closely related genetically (isozyme data) and that they are likely to be synonymous.
The previous finding that P. arecae isolates clustered closely with P. palmivora (Oudemans and Coffey 1991b, c) was supported by Mchau and Coffey (1994a), who found that the six isolates, previously identified as P. arecae, clustered with P. palmivora (ET7 and ET8). Since the isolates of both species were similar both morphologically and in isozyme patterns, they concluded that the two species are conspecific and that isolates previously designated P. arecae should be included in P. palmivora.
[[PAPER:2381|1]]
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Isolate list