
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
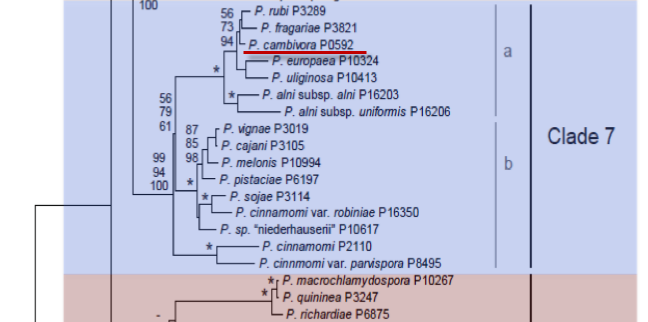
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora cambivora (Petri) Buisman 1927 (Oomycetes, Pythiales) Blepharospora cambivora Petri 1917
Blepharospora cambivora Petri 1917
Distribution: Cosmopolitan.
Substrate: Roots, stems, especially base of trunk.
Disease Note: Canker, crown and root rot, often in conjunction with other Phytophthora spp. (Waterhouse & Waterston 1966).
Host: 30 genera in 19 families, including Castanea (chestnut, Fagaceae).
Supporting Literature:
Cooke, D.E.L., Drenth, A., Duncan, J.M., Wagels, G., and Brasier, C.M. 2000. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genet. Biol. 30: 17-32
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Tucker, C.M. 1931. Taxonomy of the genus Phytophthora de Bary. Univ. Missouri Agric. Exp. Sta. Bull. 153: 1-208
Waterhouse, G.M. 1963. Key to the species of Phytophthora de Bary. Mycol. Pap. 92: 1-22
Waterhouse, G.M., and Waterston, J.M. 1966. Phytophthora cambivora. C.M.I. Descript. Pathog. Fungi Bact. 112: 1-2
Updated on Jun 05, 2006
Phytophthora cambivora (Petri) Buisman (1927) Characteristics P. cambivora is classified in group VI (Stamps et al. 1990) because it produces nonpapillate sporangia and amphigynous antheridia. Morphology is shown in Figure 1 and 2. Tabular keys are shown in Tables 4.2 and 4.3 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). Waterhouse and Waterston (1966) summarize the morphology and diseases caused.
1. Sporangia
Shapes of sporangia range from broadly ovate, subglobose, and obpyriform to ellipsoidal with well-rounded bases but are variable in shape depending on the culture media utilized. Dimensions of sporangia have been recorded as 40 to 54 x 60 to 75 µm (Petri 1917); 55 to 65 x 40 to 45 µm with maximum diameters of 85 x 60 µm (Waterhouse and Waterston 1966); 48 to 83 x 32 to 61 µm with an average of 59 to 64 x 40 to 45 µm (Chitzanidis and Kouyeas 1970) for an isolate obtained from diseased chestnut trees; 44 to 69 x 33 to 48 µm with an average diameter of 38 x 54 µm (Mircetich and Matheron 1976); 30.37 to 47.35 x 22.58 to 30.92 µm (Gerrettson-Cornell 1978); 25 to 65 x 17 to 31 µm with an average diameter of 46 x 25 µm (Suzui and Hoshino 1979); and average diameters of 50.4  10.9 x 35.2 6.7 µm for 12 isolates from several different countries (Oudemans and Coffey 1991a). Length-breadth ratios of sporangia have been recorded as 1.8 by Suzui and Hoshino (1979) and as 1.4 by Oudemans and Coffey (1991a). Sporangia proliferate internally and sometimes are nested within the original sporangium. Sporangiophores are simple, unbranched, or sympodial (Figure 1 and 2a and b).
10.9 x 35.2 6.7 µm for 12 isolates from several different countries (Oudemans and Coffey 1991a). Length-breadth ratios of sporangia have been recorded as 1.8 by Suzui and Hoshino (1979) and as 1.4 by Oudemans and Coffey (1991a). Sporangia proliferate internally and sometimes are nested within the original sporangium. Sporangiophores are simple, unbranched, or sympodial (Figure 1 and 2a and b).
2. Sex Organs
P. cambivora is normally heterothallic. Detailed morphology of the sex organs of P. cambivora shown by scanning electron microscopy and light microscopy is reported by Ho et al. (1977). Oogonia are spherical and produced terminally. The outer wall of more than 50% of the oogonia is distinctly bullate or verrucose (Figure 1 and 2d). Oogonia are normally produced abundantly only when paired with a suitable isolate of the opposite mating type of the same or different species. Some A2 isolates produce oogonia in single culture (Gerrettson-Cornell 1977). Diameters of oogonia vary from 43 µm, with a maximum of 62 µm (Waterhouse and Waterston 1966); 30 to 58 µm, average 48 µm (Chitzanidis and Kouyeas 1970); 38 to 48 µm, average 44 µm (Mircetich and Matheron 1976); average 46.3 µm (Gerrettson-Cornell 1977); average 37 µm (Suzui and Hoshino 1979); and average 40.5 5.5 µm for 12 isolates from different countries (Oudemans and Coffey 1991a). The average oogonial wall thickness is 2 µm (Waterhouse and Waterston 1966). Oospores nearly fill the oogonium (plerotic). The average reported diameters of oospores are 36 µm with a wall thickness of 3 µm (Waterhouse and Waterston 1966); 37 to 44 µm, average of 40 µm for isolates from cherry trees (Mircetich and Matheron 1976); and 35.14 to 39.42 µm for isolates obtained from eucalyptus trees (Gerrettson-Cornell 1978). The diameters of oospores of 12 isolates obtained from different countries averaged 33.8 5.6 µm (Oudemans and Coffey 1991a). The antheridia of P. cambivora are always amphigynous and are normally elongated and either unicellular or bicellular (Figures 1 and 2d); 60% of the antheridia that Ho et al. (1977) observed were bicellular. The average length of the antheridium is reported to be 25 µm with a maximum length of 35 µm (Waterhouse and Waterston 1966). Other reported diameters are 14 to 32 x 12 to 21 µm, average 19 x 16 µm (Chitzanidis and Kouyeas 1970); 19 to 38 x 15 to 21 µm (Mircetich and Matheron 1976); 16.6 to 29.4 µm, average length of 27.3 µm for the bicellular type; and 12.9 to 27.6 µm, average length 19.3 µm for the unicellular type (Ho et al. 1977).
3. Growth Temperatures
The minimum temperature for growth is 2oC, optimum is 22 to 24oC, and maximum (cessation of growth) is above 32oC.
4. Cultural Characteristics
The mycelium has coralloid hyphae often distorted into irregular shapes (Figure 1) with moderate to profuse growth of aerial mycelium. Mircetich and Matheron (1976) report that cherry isolates produce uniform and profusely fluffy cultures on cornmeal agar at 21oC. Older mycelium is often septate (Mircetich and Matheron 1976; O. K. Ribeiro, unpublished). Luo et al. (1988) report that the colony patterns of isolates varied from stellate to uniform (no discernible pattern).
5. Distinguishing Characteristics
P. cambivora has been considered to be similar to P. cinnamomi, but it differs in that it seldom produces the lightly pigmented, botryose, thin-walled hyphal swellings and chlamydospores that are produced by P. cinnamomi. Contrast the hyphal swellings illustrated in Figure 2 with those illustrated for P. cinnamomi in Figure 4.1 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). Some of the oogonia of P. cambivora are verrucose (bullate) (Figures 4.27, 13.1, and 13.2 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996); P. cinnamomi oogonia are always smooth walled. P. cambivora is serologically distinguished from P. cinnamomi and other species (Halsall 1976). Polyacrylamide gel electrophoresis of native proteins in the mycelium of several isolates of P. cambivora, obtained from various sources in the United States, showed the presence of only one protein-banding pattern that distinguished them from P. cactorum and P. syringae (Bielenin et al. 1988). Extensive isozyme analyses by starch gel electrophoresis showed that P. cambivora is distinctly separated from P. cinnamomi and P. cactorum. Eight distinct electrophoretic types were delineated in isolates of P. cambivora from different parts of the world. Isolates from Europe were of a single electrophoretic type, but five of the types were characteristic of isolates from Australia (Oudemans and Coffey 1991a).
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Isolate list