
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
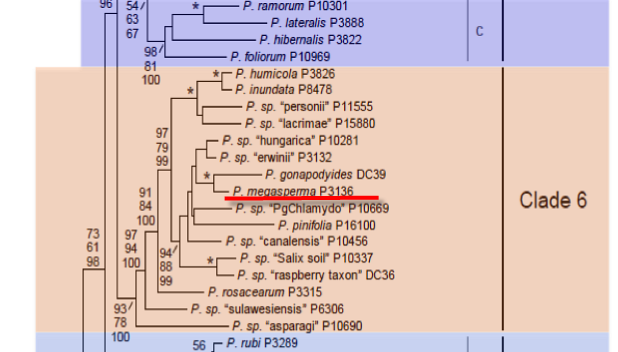
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora megasperma Drechsler 1931 (Oomycetes, Pythiales) Phytophthora megasperma var. megasperma Drechsler 1931 Note: Type variety not needed.
Phytophthora megasperma var. megasperma Drechsler 1931 Note: Type variety not needed.
= Pythiomorpha miyabeana S. Ito & Nagai 1931
Notes: Phytophthora megasperma has been divided into a number of distinct species including Phytophthora sojae, Phytophthora medicaginis, and Phytophthora trifolii. According to Cooke et al. (2000), the Phytophthora megasperma type on asparagus, the DF type on Douglas-fir, and the O group each represent distinct species as well. Varietal distinction is no longer needed.
Distribution: Cosmopolitan.
Substrate: All plant parts, depending on host.
Disease Note: Root rot, also crown rot, storage rot, seedling damping-off, fruit rot, foot rot, stem canker, tuber rot, collar rot, sudden wilt, apoplexy, stunting, chlorosis, depending on host (see Erwin & Ribeiro 1996).
Host: On Fabaceae, isolates previously referred to as P. megasperma are now considered to be three distinct species.
Supporting Literature:
Cooke, D.E.L., Drenth, A., Duncan, J.M., Wagels, G., and Brasier, C.M. 2000. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genet. Biol. 30: 17-32
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Kroon, L.P.N.M., Bakker, F.T., van den Bosch, G.B.M., Bonants, P.J.M., and Flier, W.G. 2004. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNS sequences. Fungal Genet. Biol. 41: 766-782
Waterhouse, G.M., and Waterston, J.M. 1966. Phytophthora megasperma. C.M.I. Descript. Pathog. Fungi Bact. 115: 1-2
Updated on Jun 12, 2006
P. megasperma is classified in group V (Stamps et al. 1990) because it produces nonpapillate sporangia and the antheridia are mostly paragynous. Morphology is shown in Figure 1, Figure 2 and Figure 3. Sizes of sporangia, oogonia, and oospores, proportion of paragynous and amphigynous antheridia, and growth temperatures for isolates from different hosts are given in Table 1. See Tables 4.2 and 4.3 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996) for tabular keys. The taxonomic status of P. megasperma is reviewed by Erwin (1983), E. M. Hansen and Hamm (1983), E. M. Hansen et al. (1986), E. M. Hansen (1987, 1991), E. M. Hansen and Maxwell (1991), and Förster and Coffey (1993). Classification is discussed further in Chapter 4 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996).
1. Sporangia
Sporangia are nonpapillate, ovoid, obpyriform, 35 to 60 µm long x 25 to 45 µm wide, and persistent on the stalk and proliferate internally (Drechsler 1931). See Table 1 for dimensions of sporangia of isolates from different hosts. Sporangiophores are simple or sparingly branched.
2. Hyphal Swellings
Hyphal swellings are produced in aqueous cultures and are rounded or angular and in chains or clusters.
3. Chlamydospores
Chlamydospores are not produced.
4. Sex Organs
P. megasperma biotypes are mostly homothallic. Barr (1980) and Ho (1986a) reported that a large-oospore isolate from alfalfa was heterothallic. Antheridia are irregularly spherical or ellipsoid (10 to 18 x 14 to 20 µm) and mainly paragynous; however, some amphigynous antheridia form. Oogonia are produced abundantly in host tissues and in axenic culture, are smooth and subspherical, and measure 42 to 52 µm in diameter, average 47.4 µm. Oospores are 26 to 52 µm in diameter, average 41.1 µm (Drechsler 1931) (see also Table 1).
5. Growth Temperatures
See Table 1 for growth temperature characteristics of isolates reported from various references. Drechsler (1931) did not report temperature relations. A large-oogonia isolate from hollyhock (Tompkins et al. 1936b) grew better at 25oC than at 30oC; no growth occurred at 35oC. Temperature relations of isolates from cauliflower, stock, cabbage, and cineraria were similar. In a modification of Tucker\'s (1931) key, P. megasperma was differentiated from P. syringae by growth at 28oC (Tompkins et al. 1936b).
E. M. Hansen and Maxwell (1991) report the optimum temperature for growth of Douglas fir (DF1) isolates, 27.1oC; DF2, 21.6oC; large-oogonia isolates from alfalfa (A1F2), 22.5oC; rose, 25oC; apple, 22.5oC and 20oC; grape, 20oC; juniper, 22.5oC; cherry, 25oC; pear, 25oC; poplar, 20oC; Brassica spp., 20oC; and the type culture from hollyhock (Drechsler 1931), 20oC.
[[PAPER:2373|1]]
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Isolate list