
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
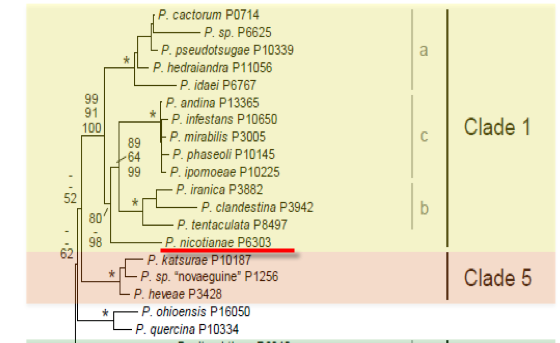
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora nicotianae Breda de Haan 1896 (Oomycetes, Pythiales) Phytophthora nicotianae var. nicotianae Breda de Haan 1896 Note: Type variety no longer needed.Phytophthora parasitica var. nicotianae (Breda de Haan) Tucker 1931
Phytophthora nicotianae var. nicotianae Breda de Haan 1896 Note: Type variety no longer needed.Phytophthora parasitica var. nicotianae (Breda de Haan) Tucker 1931
= Phytophthora allii Sawada 1915
= Phytophthora formosana Sawada 1942
= Phytophthora jatrophae Rosenbaum 1917 Note: Also described by Rutgers in 1917? Sometimes cited as Jensen, but this is an invalid name, nom. nud. (no description). Also cited as Jensen 1923 by Erwin & Ribeiro 1996.
= Phytophthora melongenae Sawada 1915 Note: See Ho & Chang 1992.
= Phytophthora parasitica Dastur 1913
Phytophthora nicotianae var. parasitica (Dastur) G.M. Waterhouse 1963
Phytophthora parasitica var. parasitica Dastur 1913
= Phytophthora parasitica var. rhei Godfrey 1923
= Phytophthora parasitica var. sesami Prasad 1957
= Phytophthora tabaci Sawada 1927 Note: See Ho & Chang 1992.
= Phytophthora terrestris Sherb. 1917 Note: Published as terrestria.
Blepharospora terrestris (Sherb.) Peyronel 1920
Notes: Varietal distinctions are no longer recognized (Erwin & Ribeiro 1996). Phytophthora parasitica var. sabdariffae Mukerjee is an invalid name, nom. nud. (Waterhouse 1963). Phytophthora nicotianae was described by Breda de Haan in 1896 based on cultures contaminated with a Pythium species. The name was never formally rejected as a mixed type under the former Art. 70 of the 1969 Code; this is no longer an option, as the article has been deleted from the current code. The name Phytophthora parasitica until recently has been frequently used for this species, and there has been some discussion of a proposal for conservation, but to date this has not happened (see discussion in Erwin & Ribeiro 1996, p. 391).
Distribution: Cosmopolitan.
Substrate: Roots, stems, trunk, leaves, fruits, pods.
Disease Note: Black shank of tobacco, damping-off, crown rot, leaf blight, fruit rot. See Erwin & Ribeiro for detailed disease notes by host.
Host: Wide host range: 255 genera in 90 families.
Supporting Literature:
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Ho, H.H., and Chang, H.S. 1992. A re-evaluation of Phytophthora species described by K. Sawada in Taiwan. Mycotaxon 43: 297-316
Kroon, L.P.N.M., Bakker, F.T., van den Bosch, G.B.M., Bonants, P.J.M., and Flier, W.G. 2004. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNS sequences. Fungal Genet. Biol. 41: 766-782
Tucker, C.M. 1931. Taxonomy of the genus Phytophthora de Bary. Univ. Missouri Agric. Exp. Sta. Bull. 153: 1-208
Waterhouse, G.M. 1963. Key to the species of Phytophthora de Bary. Mycol. Pap. 92: 1-22
Waterhouse, G.M. 1970. The genus Phytophthora de Bary. Mycol. Pap. 122: 1-59
Waterhouse, G.M., and Waterston, J.M. 1964. Phytophthora nicotianae var. parasitica. C.M.I. Descript. Pathog. Fungi Bact. 35: 1-2
Waterhouse, G.M., and Waterston, J.M. 1964. Phytophthora nicotianae var. nicotianae. C.M.I. Descript. Pathog. Fungi Bact. 34: 1-2
Updated on Jun 08, 2006
P. nicotianae is classified in group II (Stamps et al. 1990) and redescribed by G. Hall (1993). See Tables 4.2 and 4.3 for tabular keys, Appendix 4.9 for a dichotomous key (Ho 1992), Tables 51A.4 and 51A.5 for morphological comparison with P. palmivora and related species in Phytophthra Diseases Worldwide (Erwin and Ribeiro 1996), and Waterhouse and Waterston (1964d, e) for Commonwealth Mycological Institute descriptions of P. nicotianae.
1. Sporangia
Shapes of sporangia vary from ellipsoid, ovoid, pyriform, obpyriform, to spherical with a prominent papilla [see Figure 4.12C in Phytophthra Diseases Worldwide (Erwin and Ribeiro 1996)]; occasionally two papillae occur on a single sporangium. Sporangia are not caducous. Papillate sporangia are produced singly or in a loose sympodium on long stalks 100 to 595 µm in length (average 375 µm) in some isolates (Thomson and Hine 1972); sporangia are 11 to 60 µm long 20 to 45 µm wide (average 40.18 28.53 µm) with a length-breadth ratio of 1.1 to 1.7 (average 1.34). G. Hall (1993) reports highly variable sizes for different isolates and a length-breadth ratio of 1.1 to 1.3. Other ratios include 1.3 (Godfrey 1923) and 1.4 (Ashby 1928; Waterhouse 1963; Feichtenberger 1980). Sporangiophores are irregularly or sympodially branched. Waterhouse (1974c) indicated that sporangia of the variety parasitica were caducous with short stalks (2.0 µm) but the sporangia of variety nicotianae were not; however, this was not confirmed by Al-Hediathy and Tsao (1979a, b) and Trichilo and Aragaki (1982). Sporangia from isolates labeled either variety were persistent on the stalk (noncaducous). G. Hall (1993) noted that some sporangia broke off sporangiophores; but there was no septum at the breakage point, and thus they concluded that sporangia are noncaducous.
A study of the morphology of an isolate from crepe ginger, Costus speciosus (J. König) Sm., by electron and light microscopy revealed reticulate ridges on the surfaces of sporangia that had not been previously described (Khan et al. 1988).
2. Chlamydospores
Chlamydospores are usually abundant; however, G. Hall (1993) reported that only 50% of 81 isolates produced them abundantly. Chlamydospores are terminal or intercalary and 13 to 60 µm in diameter, average 28 µm. G. Hall?s (1993) neotype averaged 33 µm in diameter. Dastur (1913) reported a range of 20 to 60 µm. Hyphal swellings with thin walls were noted by G. Hall (1993).
3. Sex Organs
Most isolates are heterothallic, but some isolates form oogonia and oospores in single culture when inoculum is transferred from old cultures (Brasier 1972; Tsao et al. 1980). Isolates from vinca, Catharanthus roseus, were homothallic (Schubert and Leahy 1989). Oospores were found in stelar tissues of infected Washington Navel orange roots in Tulare County, California (Lutz and Menge 1991). Antheridia are amphigynous and spherical or oval; oogonia are smooth and spherical and range from 15 to 64 µm in diameter (average 26.8 µm). G. Hall (1993) reports 26 3.2 µm. Oospores are aplerotic; Dastur (1913) reports a range of 13 to 24 µm. Ranges of 13 to 35 µm in diameter with an average of 22.6 µm (see Table 50.1) are common. G. Hall (1993) reports a diameter of 24 3.2 µm.
Phytophthora nicotianae is the causal agent of black shank of cultivated tobacco. This disease is often initially noticed in low areas or along edges of a field. The first symptoms of black shank are usually yellowing and wilting of a few scattered plants. Black shank can be mistaken for drowning, especially if the first plants affected are in a low area. Black shank-infected plants do not recover and soon the leaves on the entire plant wilt and turn a golden yellow.
The lower stalk and root system of infected plants are usually black. The blackened area on the stalk is often sunken and extends from several inches above the soil line into the root system. Splitting the stalks of larger plants will often reveal a brown pith that is segmented into layers, called \\'discing\\'. The pith is the inner-most portion of the stalk and in healthy plants is off-white, soft and not layered. However, segmenting of the stalk pith alone should not be relied on for positive identification of black shank, since this symptom does not always occur. Also, segmenting of the pith can be caused by other factors, such as lightning.
Black shank induced discing in an infected burley tobacco stalk.

P. nicontianae infested burley tobacco field in the mountains of NC.
P. nicotianae also causes root rot on many different ornamental plants, including boxwoods.

American boxwood infected with P. nicotianae.
Tobacco: Host resistance provides an affordable and effective method of black shank control, and is based on both complete (single-gene) and partial (multigene) resistance. Single-gene resistance, derived from Nicotiana longiflora (Phl) and N. plumbaginifolia (Php), provides complete resistance to race 0 of P. nicotianae, but places high selection pressure on the pathogen for the development of races. Since the deployment of the Php gene in flue-cured tobacco, incidence of race 1 has increased dramatically, becoming the dominant race in many tobacco producing regions.
Burley tobacco plants on right having high level of resistance to race 0 of P. nicotianae (cultivar Bu 37), with susceptible Bu 21 cultivar on left.
Tobacco: Ridomil Gold is a systemic fungicide that can be used to supplement crop rotation and resistant varieties in controlling black shank. The chances of realizing economic returns from the use of Ridomil are greatest if it is used in conjunction with varieties that have at least a 4 resistance or higher rating to both Race 0 and Race 1. Ridomil should not be used for black shank control on highly susceptible burley (varieties with a resistance rating of 0-2) or dark-type varieties. The chances of obtaining economic returns are also reduced in fields heavily infested with black shank even if Ridomil is used, on varieties with levels of resistance of 2 or less. Use Ridomil Gold at 1 qt/acre, applied broadcast with a minimum of 20 gallons of water per acre and incorporated into the top 2-4 inches of soil.
For prolonged control of black shank (especially in fields that have had heavy black shank infections in the past), the following is recommended: apply Ridomil Gold at 1 pt/acre just prior to transplanting, followed by a second application at 1 pt/acre at the first cultivation, followed by a third application of 1 pt/acre at lay-by or the last cultivation. The label allows these supplemental applications only if 1 pt/acre was used at transplanting.
[[PAPER:2378|1]]
[[PAPER:2397|1]]
[[PAPER:2379|1]]
[[PAPER:2398|1]]
[[PAPER:2380|1]]
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Species page written by Kelly Ivors. Pictures provided by Kelly Ivors and Paul Shoemaker of NC State University.
Isolate list