
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
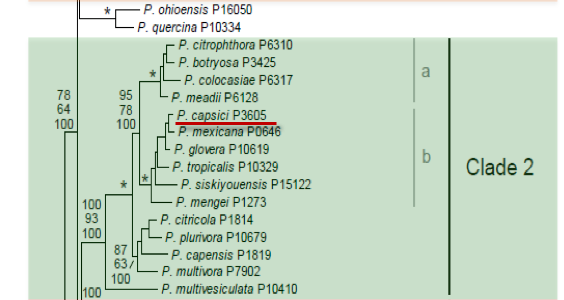
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora capsici Leonian 1922 (Oomycetes, Pythiales) Phytophthora parasitica var. capsici (Leonian) Sarej. 1936
Phytophthora parasitica var. capsici (Leonian) Sarej. 1936
= Phytophthora hydrophila Curzi 1927
Notes: Phytophthora palmivora (as P. palmivora MF4) is considered by some authors to be a partial synonym (e.g., Erwin & Ribeiro 1996). Various taxonomists including Mchau et al. (1995) have broadened the species concept of Phytophthora capsici to include a range of morphological characters and the two isozyme groups CapA or Cap 1, including -Phytophthora palmivora MF4-) and CapB or Cap 2. Aragaki & Uchida (2001) argued for a narrower species concept, referring several tropical isolates (= Cap A or Cap 1) to the new species Phytophthora tropicalis.
Distribution: Cosmopolitan. Tropical species may be Phytophthora tropicalis.
Substrate: Roots, fruits, stems, seedlings, pods, cotton bolls.
Disease Note: Fruit, stem, and root rot. Also seedling damping-off, leaf wilt.
Host: Phytophthora capsici sensu lato infects 51 genera in 28 families, including Capsicum annuum (peppers), Lycopersicon esculentum (tomatoes), and other Solanaceae. Aragaki & Uchida 2001 refer isolates from non-Capsicum hosts to P. tropicalis, including isolates from Macadamia spp., Theobroma cacao (cacao), and other tropical crop species.
Supporting Literature:
Aragaki, M., and Uchida, J.Y. 2001. Morphological distinctions between Phytophthora capsici and P. tropicalis sp. nov. Mycologia 93: 137-145
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Mchau, G.R.A., and Coffey, M.D. 1995. Evidence for the existence of two subpopulations in Phytophthora capsici and a redescription of the species. Mycol. Res. 99: 89-102
Stamps, D.J. 1985. Phytophthora capsici. C.M.I. Descript. Pathog. Fungi Bact. 836: 1
Tucker, C.M. 1931. Taxonomy of the genus Phytophthora de Bary. Univ. Missouri Agric. Exp. Sta. Bull. 153: 1-208
Waterhouse, G.M. 1963. Key to the species of Phytophthora de Bary. Mycol. Pap. 92: 1-22
Updated on Jun 05, 2006
Isozyme analysis of 84 isolates from a world collection indicates that P. capsici is a genetically complex species containing three subgroups (Oudemans and Coffey 1991b). One subgroup, CAP1, contains a majority of the isolates from annual solanaceous and cucurbit hosts as well as some isolates from black pepper and cocoa that had been previously described as P. palmivora MF4 (Kaosiri and Zentmyer 1980). A second subgroup, CAP2, contains isolates mainly from tropical crops such as black pepper, cocoa, papaya, macadamia, and rubber and isolates from Hawaii tentatively proposed as P. tropicalis by Uchida and Aragaki (1989) and Aragaki and Uchida (1992). The third subgroup, CAP3, with the least genetic diversity, includes cocoa isolates from Brazil. Isolates previously designated P. palmivora MF4 occurred in all three subgroups. Förster and Coffey (1991), in a summary of data from Förster et al. (1990a) that involved mtDNA RFLP patterns, also noted that P. palmivora MF4 grouped with isolates of P. capsici from C. annuum (pepper). Mchau and Coffey (1995), utilizing 113 isolates of P. capsici, most of which were included in the report of Oudemans and Coffey (1991b), conducted further isozyme analyses. By use of 15 enzymes, the isolates were shown to be separable into two subgroups, CAPA and CAPB. Each subgroup contained isolates from a wide range of hosts and geographical locations. However, some members of each subgroup varied in morphology. This supports the redescription by Tsao and Alizadeh (1988) that consolidates these types under P. capsici. A subsequent redescription, which adds the genetic isozyme data to morphological data, is given by Mchau and Coffey (1995). Blaha (1990) differentiated P. palmivora, P. megakarya, and P. capsici by use of two isozyme systems. Hwang et al. (1991) distinguished four groups from P. capsici pepper isolates on the basis of mtDNA RFLP patterns. Spore types of P. capsici are shown in Figure 1 and morphology is shown in Figure 2. P. capsici is classified in group II (Stamps et al. 1990). See Tables 4.2 and 4.3 for tabular keys, Appendix 4.9 for a dichotomous key (Ho 1992; Ho et al. 1995) in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996), and Stamps (1985f) for a summary. See also Tables 51A.4 and 51A.5 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996) for morphological comparisons with P. palmivora and related species.
1. Sporangia
The morphology of sporangia is shown in Figures 4.16, 4.17A, B, and E, in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996), and Figure 1 and Figure 2. Sporangia are mostly papillate, but in some cases they appear to be semipapillate (apical thickening ranges from 1.4 to 9.2 µm). Occasionally sporangia have two or three apices. Sporangial shapes are influenced by light and other cultural conditions (Tsao and Alizadeh 1988; Tsao 1991) and range from subspherical, ovoid, obovoid, ellipsoid, fusiform, pyriform, to distorted shapes. Sporangia are predominately tapered at the base and are caducous with long pedicels varying in length from 35 to 138 µm (Mchau and Coffey 1995). Length-breadth ratios in different reports are 1.70:1 (Frezzi 1950); 1.72:1 (Ershad 1971); 1.57 to 2.19, average 1.76:1 in agar and 1.52 to 2.10, average 1.73:1 in water (Kröber 1985); in light 1.73 and in dark 1.27 (Tsao and Alizadeh 1988); and 1.4 to 1.8 (range for 24 isolates incubated in water under light) (Ristaino 1990). Mchau and Coffey (1995) report that sporangia are extremely variable in shape; shapes include ellipsoid, spherical, subspherical, broadly ovoid, obturbinate, obovoid, fusiform, and pyriform. Dimensions (length breadth) varied from 32.8 to 65.8 17.4 to 38.7 µm. Length-breadth ratios varied from 1.3:1 to 2.1:1. The larger length-breadth ratio was more typical of the CAPB subgroup and the smaller of the CAPA subgroup. See Table 1 for sporangial dimensions. Sporangiophores that are formed under light are irregularly branched, and sympodia form only in water. Black pepper and cocoa (P. palmivora MF4) isolates produce sporangia on sporangiophores that are characteristically umbellate Figures 4.5 and 4.16B-F in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). Caducous sporangia with long stalks are shown in Figures 4.17A and B and 14.3. in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). Stalk lengths varied among isolates from pepper and cucurbit hosts from 37.5 to 98.6 µm for pepper isolates and 31.5 to 85.3 for cucurbit isolates (Ristaino 1990). Most isolates of subgroup CAPA produced rounded ellipsoid sporangia, some of which had more than one papilla. The majority of the isolates of subgroup CAPB produced ellipsoid-lanceolate sporangia. Since ellipsoid-lanceolate sporangia were more characteristic of CAPB isolates than of CAPA isolates, the length-breadth ratio for CAPB was higher than that for CAPA. Sporangia in CAPB were produced on umbellate sporangiophores. However, these morphological traits were not characteristic of all isolates within the subgroup. Pedicel length varied from 34.7 to 138 µm, but the mean value was significantly higher for CAPB. Since both ovoid and ellipsoid-lanceolate sporangia occurred within each subgroup, Mchau and Coffey (1995) concluded that the elongated sporangial morphology that was suggested by Uchida and Aragaki (1989) and Aragaki and Uchida (1992) to be typical of a proposed new species, P. tropicalis, was not justified.
2. Chlamydospores
Chlamydospores of P. capsici isolates from pepper (C. annuum) are rare in culture according to Tucker (1931). Pepper or cucurbit isolates did not form chlamydospores (Ristaino 1990); however, chlamydospores were reported on some pepper isolates from Iran (Ershad 1971). Chlamydospores are abundant in isolates from black pepper (Piper niger), cocoa, and macadamia (Alizadeh and Tsao 1985b; Uchida and Aragaki 1985; Tsao 1991). Diameters are 28 to 29 µm with wall thickness of 2.4 to 2.7 µm (Tsao 1991; Figure 4.25). Mchau and Coffey (1995) report chlamydospore production exclusively in a population of isolates from cacao, black pepper, and other hosts (but not from Capsicum pepper) that were differentiated isozymically into the subgroup CAPB. Chlamydospores were terminal or intercalary and varied from 22 to 52 µm in diameter. Production of chlamydospores in some isolates is conditioned to some extent by the choice of cultural methods (Uchida and Aragaki 1985). When mycelial mats were grown for 5 days on clarified V8 juice neutralized with calcium carbonate and then submerged in sterile distilled water for 4 to 8 weeks and incubated in the dark, 20 of 29 isolates of P. capsici produced chlamydospores; however, isolates from solanaceous hosts, except for three isolates from eggplant, did not produce chlamydospores. A 50% papaya juice broth previously used to induce oospores of P. palmivora (Kadooka and Ko 1973) did not induce chlamydospores of any P. capsici isolate (Uchida and Aragaki 1985). Walls were 1.0 to 1.5 µm thick. Hyphal swellings are occasionally produced by some isolates in aqueous cultures.
3. Sex Organs
P. capsici is predominately heterothallic; however, oospores have been noted to form in single culture in media containing the fungicide chloroneb (Noon and Hickman 1974) after wounding of the mycelium by exposing cultures to the fungus Trichoderma (Brasier 1975a, b, 1978) and in single A1 cultures (Kamjaipai and Ui 1978). Oospores formed in roots and stems of C. annuum inoculated with A1 and A2 mating types (Matsuoka and Ansani 1984). Both A1 and A2 mating types are isolated from pepper and cucurbit plants in the field, but in general A1 or A2 mating types occur singly and seldom together on a plant (Ristaino 1990). Antheridia are amphigynous. Diameters given in several reports include 12 to 21 x 12 to 17 µm (Frezzi 1950); 17 x 15 µm (Stamps 1985); 9 to 20 x 8 to 17 µm, average 14.1 13.5 µm (Kröber 1985); 12 to 17 14 to 16 µm, average 14 x 15 µm (Tsao 1991). Oogonia are spherical or subspherical and hyaline to brown. Dimensions of oogonia from different hosts vary from 23 to 50 µm. Oospores are predominately plerotic with a wall thickness of 2 to 6 µm (Tsao 1991). Mean diameters of oospores from several pepper isolates ranged from 23.7 to 34.9 µm; mean diameters of several cucurbit isolates ranged from 27.8 to 34.2 µm (Ristaino 1990). Dimensions of oospores from various hosts are given in Table 1.
5. Growth Temperatures
Minimum temperature for growth is 10oC, optimum is 28oC, and maximum is >35oC (Stamps 1985). Several reports show different data for minimum, optimum, and maximum temperatures: 7.5, 30, and 35oC (Kröber 1985); 6 to 9, 27 to 30, and 33 to 39oC (Tsao 1991); and 12, 28, and 32oC (Leu and Kao 1981). Isolates from CAP1 with the exception of three isolates grew well at 35oC. Four isolates of CAP2 grew poorly at 35oC, but the remaining isolates grew well (0.7?.3 mm per day). Isolates from CAP3 grew poorly or not at all at 35oC. Optimum temperatures for growth among all isolates varied from 24 to 33oC (Mchau and Coffey 1995).
6. Distinguishing Characteristics
P. capsici is distinguished from other species in group II by its production of long pedicels on caducous sporangia with shapes varying from nearly spherical to elongated with a tapering base. P. parasitica produces more rounded sporangia that are noncaducous; P. palmivora produces ovoid sporangia that are caducous but have short pedicels.
P. capsici, as well as other Phytophthora spp., produces a wide variety of symptoms on mature plants that vary according to host.
Capsicum annuum (pepper)
Roots, stems, foliage, and fruit of mature pepper plants are susceptible. Although infection occurs at any height on stems, it is most common at the soil line, and starts as a dark, water-soaked area. Stem lesions become dark brown to black and result in girdling and plant death. Infected roots become dark brown and disintegrate. Leaf spots start off small and are irregular to round, and water-soaked. With time, foliar spots enlarge, turn a light tan, and may crack. Rapid blighting of new leaves and the entire emerging shoot may take place. Pepper fruit can become infected through the fruit stalk. Fruit rot appears as dark green, water-soaked areas that become covered with a white to gray, greasy looking mold. Infected fruit dries, becomes shrunken, wrinkled, and brown, and remains attached to the stem.
Lycopersicon esculentum (tomato)
P. capsici can cause crown infections, leaf spot, and foliar blight in tomato transplants, which are most susceptible within the first four weeks after planting in the field. Diseased crowns become brown and soft and the plant may wilt and topple over. However the most common symptom is fruit rot, also known as buckeye rot. Uninjured fruit of any age may be infected. Rot is most prevalent where soil comes in contact fruit; fruit rot begins as dark, water-soaked spots. The spot rapidly expands during warm weather and covers 50% or more of the fruit surface with a brown, watery discoloration that may assume the appearance of concentric rings. Over time and under humid conditions infected fruit may be covered with white fungal growth and rot entirely following invasion by secondary microorganisms.
Cucurbita maxima (squash and pumpkin)
Cucurbit species are highly susceptible to Phytophthora foliar blight and fruit rot. Early foliar symptoms include rapidly expanding, irregular, water-soaked lesions in leaves. Dieback of shoot tips, wilting, shoot rot, and plant death quickly follow initial infection. Sunken, dark, water-soaked areas appear on infected fruit, and are rapidly covered by white fungal growth. Given optimal warm, wet weather, P. capsici can devastate entire cucurbit plantings in a matter of days.
[[PAPER:2316|1]]
[[PAPER:2279|1]]
[[PAPER:2317|1]]
[[PAPER:2318|1]]
[[PAPER:2319|1]]
[[PAPER:2320|1]]
[[PAPER:2321|1]]
[[PAPER:2322|1]]
[[PAPER:2323|1]]
[[PAPER:2324|1]]
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Species page was written by Kelly Ivors; Images came from Kelly Ivors and Kurt Lamour.