
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
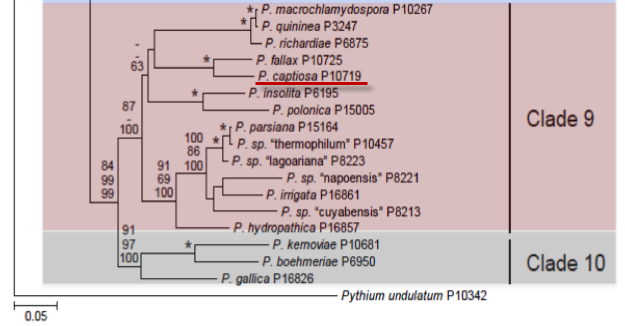
Phytophthora captiosa M. A. Dick & Dobbie was recovered from plantation Eucalyptus trees in New Zealand exhibiting a crown disease. Phylogenetically it is in clade 9 closely related to P. fallax.
1. Sporangia
Nonpapillate, non-caducous, ovoid sporangia are sometimes produced on host petiole material floated in soil water. Of the 16 isolates examined only four developed sporangia in sufficient numbers to be included in this study. Sporangia average 34 ± 4.5 x 25.5 ± 4.5 µm with isolate means of 30.5–40.5 x 20.5–29.5 µm (holotype 33.5 ± 4.5 x 29 ± 4.5 µm). Length to breath ratio is 1.2–1.5. Sporangial proliferation is both internal and external from the base of the existing sporangium.
2. Chlamydospores
No hyphal swellings or chlamydospores observed.
3. Sex Organs
Phytophthora captiosa is homothallic. Gametangia often develop in the host tissue. Eight of the 16 isolates examined produced gametangia on CA after 30 d. Oogonia on CA are spherical, averaging 33.5 ± 5.5 µm, with isolate means ranging from 30–41.5 µm (holotype 34 ± 3.5 µm). Oogonia often exhibiting a tapered base, generally turning pale brown with age. Oogonial stalk often two celled. Antheridia on CA are amphigynous, ranging from cylindrical to conical; often with coiled hyphae at the base, occasionally two celled, the bottom cell about one third the size of the top cell, averaging 19.5 ± 3 x 14 ± 2.5 µm (holotype 19 ± 4 x 11 ± 1.5 µm). Oospores are initially plerotic, occasionally becoming aplerotic with age, and averaging 29.5 ± 4 µm diameter, with isolate means ranging from 23.5– 34 µm (holotype 31 ± 3 µm). Oospore wall averaging 1.5 ± 0.5 µm thick; occasionally up to 3 µm. Oogonia formed in host tissue are brown, and slightly smaller than those produced on CA, averaging 27.5 ± 1.5 µm diameter. Antheridia in host tissue mostly single celled, brown, globose to only slightly cylindrical and averaging 14 ± 2.5 x 13.5 ± 3 µm.
4. Growth Temperatures
Grows moderately slowly on CA at 20° C in the dark, approximately 5.3 ± 0.5 mm/d (the range for five isolates was 4.6–5.9 mm/d, holotype approximately 5.6mm/d). On CA minimal growth was seen at 2° C, optimum ca 25° C, and a minuscule amount of growth was seen at 30° C. Growth ceased at 32° C.
5. Growth Characteristics in Culture
Forms stellate to rosaceous, appressed to fluffy colonies after 10 d on CA in darkness.
6. Distinguishing Characteristics
Being homothallic with amphigynous antheridia and non-papillate sporangia, P. captiosa falls in Group VI of the traditional morphological classification system while its ITS rDNA sequence indicates a closer phylogenetic relatedness to P. fallax and to Phytophthora species in ITS clades 9 and 10 (including P. insolita, P. macrochlamydospora, and P. richardiae). P. captiosa can be distinguished culturally from similar group VI species by its combination of homothallism, sometimes rather conical amphigynous antheridia, sporangial dimensions, the absence of hyphal swellings and chlamydospores, and its growth–temperature relationships. It has some similarities to P. cajani and P. melonis. However, the first formed sporangia of P. cajani are larger than those of P. captiosa at 60 X 32 µm, and later formed sporangia of P. cajani are more irregular in shape. Also P. cajani is reportedly host specific to Cajanus cajan. No growth temperature data for P. cajani were available for comparison between these two species. P. melonis is most probably heterothallic. A factor that can clearly distinguish P. melonis from P. captiosa is its minimum and optimum and maximum temperatures for growth of 9° C, 28–32° C and 37° C compared with 0–5° C, 25° C and 32° C, respectively, for P. captiosa.
Among the clades 9 and 10 Phytophthora species P. captiosa is most similar to P. richardiae. However, P. captiosa does not produce paragynous antheridia and has a lower minimum growth temperature (2° C) than P. richardiae, which does not grow below 10° C. Morphologically, P. captiosa can be distinguished from P. insolita by its formation of antheridia, as P. insolita has oogonia devoid of antheridia. It can be distinguished from P. macrochlamydospora by the absence of chlamydospores.
P. captiosa causes a crown disease of Eucalyptus botryoides and E. saligna, symptoms include leaf spots, petiole infection and twig and small branch lesions.
Dick, M. A., Dobbie, K., Cooke, D. E. L., Brasier, C. M. 2006. Phytophthora captiosa sp. nov. and P. fallax sp. nov. causing crown dieback of Eucalyptus in New Zealand. Mycol. Res. 110:393-404.
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS. This species page was adapted from Dick et al. (2006)
Isolate list