
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
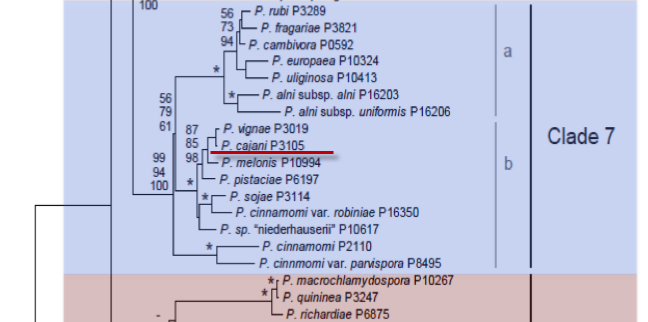
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora cajani K.S. Amin, Baldev & F.J. Williams 1978 (Oomycetes, Pythiales)
= Phytophthora drechsleri f. sp. cajani Kannaiyan 1980
= Phytophthora drechsleri var. cajani M. Pal, Grewel & A.K. Sarbhoy 1970
Notes: Cooke (2000) found no support for treating this as a variety or f. sp. of Phytophthora drechsleri. Amin et al. (1978) described P. cajani without reference to P. drechsleri var. cajani. See also Erwin & Ribeiro (1996) for discussion of taxonomy.
Distribution: Asia (India).
Substrate: Stems, also on branches, petioles, leaves. Roots are not attacked.
Disease Note: Stem rot and blight.
Host: Cajanus cajani (Fabaceae).
Supporting Literature:
Cooke, D.E.L., Drenth, A., Duncan, J.M., Wagels, G., and Brasier, C.M. 2000. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genet. Biol. 30: 17-32
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Updated on Jun 06, 2006
P. cajani is classified in group VI by Stamps et al. (1990). See Tables 4.2 and 4.3 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996) for tabular keys. Characteristics given here are from the description of P. cajani by Amin et al. (1978).
Sporangia
Sporangia form readily on infected pigeon pea stems incubated in distilled water for 36 to 48 h. Sporangia are ovoid, obpyriform, and persistent. The apical thickening is subhemispherical and 1.5 µm deep ("slightly conspicuous"). First- formed sporangia average 60 x 32 µm with a length-breadth ratio of 1.7:1.0. Sporangia formed later are smaller and more irregular in shape. The diameter at the exit pore averages 9.4 µm, a feature that Amin et al. (1978) considered to be unique.
Hyphal Swellings
Hyphal swellings have not been described.
Sex Organs
Oogonia formed in 2 to 3 weeks when P. cajani was grown on lima bean, oatmeal, or cornmeal agar and on autoclaved leaves of Cynodon dactylon (L.) Pers. incubated in distilled water at 30oC for 6 to 7 days (homothallic). Oogonia are 22 to 40 µm in diameter (average is 33.8 µm on lima bean agar, 40.2 µm on oatmeal agar, and 35.5 µm on cornmeal agar). Antheridia are amphigynous and 15 to 16 µm in diameter. Oospores are spherical, smooth, plerotic, and 20 to 37 µm in diameter.
Growth Temperatures
Growth temperatures were not reported by Amin et al. (1978).
Distinguishing Characteristics
Amin et al. (1978) report that sporangium size is close to that of P. cinnamomi, P. cambivora, P. erythroseptica var. pisi, and P. oryzae (P. japonica), but P. cajani differs from P. cinnamomi by not producing botryose hyphal swellings or chlamydospores, from P. cambivora by production of narrower sporangia and no protuberances on the oogonial wall, from P. erythroseptica var. pisi by "lacking a medial sporangium construction," and from P. oryzae by production of single sporangiophores that bear a single terminal sporangium and by production of sex organs in single culture (homothallic). Amin et al. (1978) recognized that P. cajani resembled P. drechsleri but noted that P. cajani produced larger sporangia and undifferentiated sporangiophores. They also considered that the homothallic nature of P. cajani differentiated it from P. drechsleri, which is considered to be heterothallic according to Savage et al. (1968). As stated above, homothallism has been noted in several reports on P. drechsleri (Kannaiyan et al. 1980). Our conclusion, based on Kannaiyan et al. (1980), is that P. cajani is probably the same as P. drechsleri var. cajani and P. drechsleri f. sp. cajani. The forma specialis designation is supported by its host specificity to pigeon pea and the closely related wild Atylosia sp. and the lack of pathogenicity to other hosts that are susceptible to P. drechsleri. The data of Mills et al. (1991) strongly suggest that the pigeon pea blight pathogen is significantly different from other P. drechsleri isolates and if re-described could be renamed P. cajani.
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Isolate list