
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
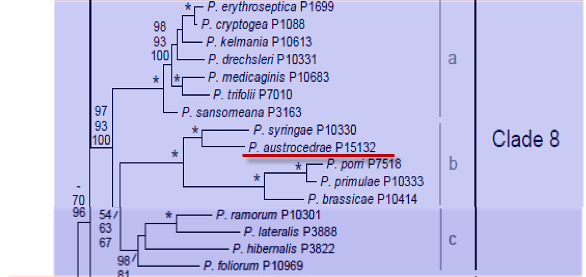
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora austrocedri Gresl. & E.M. Hansen 2007 (Oomycetes, Pythiales)
Variant spelling Phytophthora austrocedrae Gresl. & E.M. Hansen 2007
Distribution: Argentina.
Substrate: Isolated from necrotic lesions of stems and roots.
Disease Note: Associated with 'mal del cipres' disease.
Host: Austrocedrus chilensis (Cupressaceae).
Supporting Literature:
Greslebin, A.G., Hansen, E.M., and Sutton, W. 2007. Phytophthora austrocedrae sp. nov., a new species associated with Austrocedrus chilensis mortality in Patagonia (Argentina). Mycol. Res. 111: 308-316.
Updated on Nov 17, 2009
Phytophthora austrocedrae Gresl. & E. M. Hansen was isolated from necrotic lesions of stem and roots
of Austrocedrus chilensis in Argentina. Phylogenetically is closely related to P. syringae in clade 8b.
1. Sporangia
Hyphal swellings usually formed in solid and liquid media, but were more abundant in the former. Swellings were globose to subglobose and catennulated, sometimes with distorted shapes. Sporangiophores were mostly simple, 3–11 µm diameter, frequently with hyphal swellings. Sporangia were borne terminally on mostly unbranched sporangiophores. They were ovoid, obpiriform, limoniform or ellipsoid; semi-papillate, papilla 1–3(–5) µm thick, non-papillate sporangia were infrequently observed. They measured in average 50 ± 12 X 36 ± 7 µm (range 22–83 X 15–58 µm) length:breadth ratio average 1.4 ± 0.2 (range 1.1–2) and infrequently had distorted shapes. Sporangia with hyphal projections and lateral attachment of the sporangiophore were frequently observed in all isolates. The abundance of sporangia in water culture (soil extract or river water) was variable. Sporangia were not observed in solid media.
2. Chlamydospores
3. Sex Organs
Oogonia formed in single- strain culture in PAR, NAR, V8A, TA (tomato agar), CMA, CMAb (CMA plus β-sitosterol), PDAb. Oogonia were usually formed in selective media after about 20 days. They usually formed more quickly and were more abundant on selective media than on media without antibiotics. Oogonia were globose or nearly so, on average 38.5 ± 7 X 39 ± 6 µm diameter (range 22–56 µm), with hyaline to light brown, smooth walls. Oospores were globose, in average 31 ± 6 µm diam (range 17–48 µm), hyaline, with smooth walls 1–2(–3) µm thick. Antheridia were amphigynous, hyaline, one celled, in average 18 ± 3.5 X 14 ± 2 µm (range 10–30 X 8–20 µm).
4. Growth Temperatures
Growth was very slow and favored by cool temperatures. Optimum temperature was 17.5° C, with no growth at 25° C. Maximum radial growth rate of eight isolates on V-8 agar ranged from 1–1.8mm/ d.
5. Growth Characteristics in Culture
Colony in V8A, TA and TAb was uniform, without growth pattern, cottony, dome-shaped in the centre and appressed or mostly submerged at the margins. In CMA and CMAb the colony was appressed, with little or no aerial mycelium; the submerged mycelia showed an arachnoid pattern. In PDA the colony was uniform, without growth pattern, densely felty to woolly, with abundant and dense aerial mycelium and defined margins.
Growth rate on CMA and PDA was affected by β-sitosterol, being faster when β-sitosterol was added to the media. Hyphae were (3–)4–8 µm in diam. Morphology of hyphae varied according to the growth medium. This variation seemed to be related to the amount of β-sitosterol in the media. In V8A, TA, TAb, PDA and PDAb hyphae of the margin of the colony were mostly straight (or nearly so), with sparse, long branches at nearly right angles. Behind the margins, hyphae usually produced short, digitiform branches. Hyphae of the central area of the colony were mostly irregular in diam. with lateral swellings; these swellings were also observed, though less frequently, in hyphae at the colony margin. In unamended CMA, hyphae of the advancing zone looked tortuous, sometimes with lateral swellings. They were ramified at more or less right angles, with each long branch with many very short, digitiform branches. In some cases branches originated from the same point on the main hyphae and were radiate. The tortuous aspect of the hyphae and the short, digitiform branches were not present in hyphae from the advancing zone on CMA media when it was amended with β-sitosterol (30 mg/ l). Hyphae with thickened walls were observed, though infrequently, on T, TAb and PDAb after four weeks.
6. Distinguishing Characteristics
This species is characterized by the combination of a very slow growth rate, semipapillate, non-caducous and non-proliferating sporangia, oogonia with amphigynous antheridia formed in single culture, and low (17.5° C) optimal temperature for growth. In addition, the morphology of mycelia is often characteristic and may be useful for the identification of the species in combination with the other characters. It can be distinguished from its closest relative Phytophthora syringae by colony pattern and antheridia. P. syringae has a petaloid colony pattern in V8A, TA and PDA, and oogonia with paragynous antheridia that, in isolates from Patagonia, are usually formed only in media amended with oil and at temperatures below 12° C. Sporangia and hyphal swellings are very similar in both species, but although direct germination of sporangia is common in Phytophthora syringae, it was not observed in P. austrocedrae. Cardinal and optimal temperatures of growth of P. syringae are similar to those of. P. austrocedrae.
It is homothallic, readily distinguishing it from the heterothallic species in group IV (P. colocasiae, P. infestans, and P. mirabilis) and from P. macrochlamydospora. Noncaducous sporangia, hyphal swellings readily formed in water and solid culture and slow growth rate at optimal temperature differentiate this species from homothallic group IV species like P. hibernalis, P. ilicis, P. phaseoli and P. psychrophila. While there are similarities between P. austrocedrae and P. psychrophila (i.e. colony pattern, sporangia, sex organs, cardinal and optimal temperatures for growth) there are clear differences in their ITS rDNA sequences. P. psychrophila belongs to clade 3, whereas P. austrocedrae belongs to clade 8. In culture they can by differentiated by growth rate (P. psychrophila 4.2 mm d/1 in V8A) and by lack of hyphal swellings in P. psychrophila.
Phytophthora austrocedrae isolated from necrotic lesions of stem and roots of Austrocedrus chilensis. Isolation from diseased trees was successful when plates were incubated at 17° C, and unsuccessful at temperatures of 20° C or above. P. austrocedrae isolation frequency was higher on PAR and NAR than BARP and PARNBP. CMA was as successful as PAR and NAR for laboratory cultures. No differences were detected between isolations made in the field by direct plating of necrotic tissues into selective media and those made in the laboratory after washing of tissues with running tap water for 24–48 h showing that phenolyc and other water-soluble compounds did not inhibit mycelial growth. In all media, first growth from bark pieces was not visible until 10 d or more after plating.
Greslebin, A.G, Hansen, E. M., Sutton, W. 2007. Phytophthora austrocedrae sp. nov., a new species associated with Austrocedrus chilensis mortality in Patagonia (Argentina). Mycol. Res. 111:308-316.
This species page was adapted from Greslebin et al. (2007).
Isolate list