
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
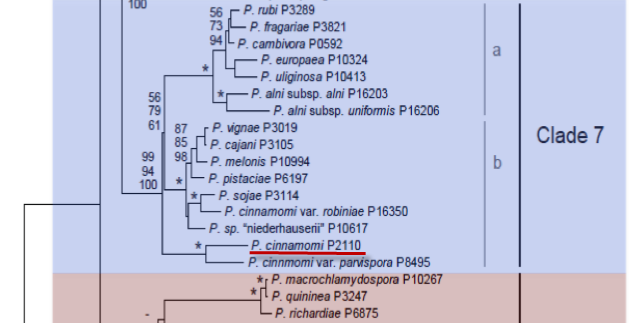
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora cinnamomi var. cinnamomi Rands 1922 (Oomycetes, Pythiales) Phytophthora cinnamomi Rands 1922 Note: See type variety.
Phytophthora cinnamomi Rands 1922 Note: See type variety.
Notes: Previously considered a synonym of Phytophthora cambivora, but now recognized as distinct (Waterhouse 1963).
Distribution: Cosmopolitan. P. cinnamomi occurs worldwide with the predominant mating type being A2.
Substrate: Roots, stems, heartwood.
Disease Note: Root rot, heart rot, wilt. Causes ink disease of Chestnut in conjunction with Phytophthora cambivora. A serious pathogen of hardwood forests and various crop species.
Host: 266 genera in 90 families, commonly hardwood trees.
Supporting Literature:
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Ho, H.H., and Chang, H.S. 1992. A re-evaluation of Phytophthora species described by K. Sawada in Taiwan. Mycotaxon 43: 297-316
Kroon, L.P.N.M., Bakker, F.T., van den Bosch, G.B.M., Bonants, P.J.M., and Flier, W.G. 2004. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNS sequences. Fungal Genet. Biol. 41: 766-782
Waterhouse, G.M. 1963. Key to the species of Phytophthora de Bary. Mycol. Pap. 92: 1-22
Waterhouse, G.M. 1970. The genus Phytophthora de Bary. Mycol. Pap. 122: 1-59
Waterhouse, G.M., and Waterston, J.M. 1966. Phytophthora cinnamomi. C.M.I. Descript. Pathog. Fungi Bact. 113: 1-2
Updated on Jun 06, 2006
See Figure 1 for morphology. The following description is reproduced by permission of the Journal of Phytopathology, courtesy Kröber and Marwitz (1993). Kröber and Marwitz (1993) state that P. cinnamomi var. parvispora differs from P. cinnamomi because it produces smaller chlamydospores (27.1 μm in diameter) than P. cinnamomi (41 μm) (Rands 1922). They also state that sporangia are smaller, a statement that is borne out by sporangial size data shown in Table 15A.1 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). Nonetheless, since sporangial size is influenced by so many factors (see Chapter 3 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996), the size of P. cinnamomi var. parvispora sporangia might simply be within the range of normal variability. A higher maximum temperature for growth of P. cinnamomi var. parvispora (36-37oC) is also cited as a difference; however, Table 4 in Zentmyer (1980) lists maximum temperatures from various reports on P. cinnamomi as 30-36oC. Since DNA RFLP technology has become such a useful tool in determining genetic differences between species, it will be of interest to determine how P. cinnamomi var. parvispora compares with other isolates of P. cinnamomi and/or P. cryptogea that Kröber and Marwitz (1993) say it resembles. Oudemans and Coffey (1991a) showed that isozyme patterns of P. cinnamomi isolates from a worldwide collection differed distinctly from those of P. cryptogea. The various isolates of P. cinnamomi were relatively uniform; however, eight electrophoretic types could be distinguished.
1. Sporangia
Hyphal swellings abundant; mostly side twigs coralloidly blown up, sometimes allantoid or tailshaped, in part several times constricted, or spherical. Zoosporangia without papilla; ovoid, ellipsoidal, obpyriform or rarely irregularly shaped; single or several in succession with internal proliferation, sometimes nest-shaped; 14-83 13-43 (36.9 25.5) μm, l/b = 1.45/1.
2. Chlamydospores
Chlamydospores abundant; spherical; terminal; intercalary, rarely lateral, sometimes in clusters on short side stalks; thin-walled; 16-43 (27.1) μm.
3. Sex organs
Generative organs only when complementary strains grown together in intraspecific mating or with P. cinnamomi or P. cryptogea in interspecific mating. Oogonia in intraspecific mating culture abundant; spherical; 23-41 (31.9) μm. Antheridia 1 per oogonium; amphigynous; spherical, broad ellipsoidal; relatively large. Oospores spherical; with a moderately thick wall; hyaline; generally almost plerotic; 22-36 (28.1) μm.
4. Growth temperature
Minimum 10-11, optimum 26-32, maximum 36-37oC; growth rate 10-14 mm/24 hr at optimum temperatures.
5. Growth characteristics in culture media
Mycelium on carrot agar hyaline; in strands; transparent; only a few downy airmycelia.
6. Distinguishing characteristics
Isolated from stem bases of Beaucarnea spec.; Germany (in greenhouses), 1990. P. cinnamomi var. parvispora is in [sic] some extent closely connected with P. cryptogea Pethybridge et Lafferty and P. drechsleri Tucker but nearest with P. cinnamomi Rands. But it is clearly distinct even from this fungus by considerably smaller chlamydospores and zoosporangia and it has a higher demand of temperature.
FRASER FIR ROOT ROT. Phytophthora root rot, caused by several Phytophthora spp., including P. cinnamomi, has been associated with significant damage to Fraser fir (Abies fraseri) in the southern Appalachian region since the 1960s. Phytophthora species represent a serious limitation to the U.S. Christmas tree market, a 3 million industry in 2003 (which includes Fraser fir, Douglas fir, Scotch pine, white pine, and other species). However annual losses due to Phytophthora are estimated at -7 million and long term viability of the industry is now in doubt (Frampton et al., 2003). Several species of Phytophthora, including P. cinnamomi, P. citricola (Shew & Benson, 1981), P. cactorum (Benson & Grand, 2000), and P. drechleri (Benson et al., 1976) have been isolated from symptomatic roots and surrounding soils, although P. cinnamomi accounts for most of the disease incidence in Fraser fir (Benson & Grand, 2000). The disease is correlated with abnormally high soil moisture promoted by frequent precipitation, flooding and poor soil drainage (Kenerley et al., 1984). P. cinnamomi can spread by use of contaminated equipment, with infected nursery stock, or with water runoff from nearby infested sites. Transplanting infected nursery seedlings represents a major contribution to disease incidence in the field, yet there are no current standard methods for certifying seedlings as pathogen free. Benson & Grand (2000) reported that 2% of transplants judged healthy based on visual inspection were infected with P. cinnamomi. Although this percentage appears small, it can translate to approximately 8 infected trees per hectare in a standard 5 ft by 5 ft spacing. Trees down-slope of introduction points can become infected from surface flow during rain events. Infected seedlings not exhibiting symptoms at field transplanting may later succumb to Phytophthora during periods that are favorable for disease development, resulting in further economic loss to the grower and infestation of previously uncontaminated sites. After disease develops in the field, growers have few options beyond harvesting as soon as possible as the pathogen can survive extended periods of time as chlamydospores in the soil, in pieces of organic matter, or in roots of fir seedlings and trees. Due to this persistence, once the pathogen becomes established on a site, the area is often no longer used for Fraser fir production. Aboveground symptoms of the disease, such as yellow-green needles and dead branches, are not obvious until the roots are heavily colonized, after which death of the tree usually ensues.
[[PAPER:2239|1]]
[[PAPER:2326|1]]
[[PAPER:2327|1]]
[[PAPER:2328|1]]
[[PAPER:2329|1]]
[[PAPER:2238|1]]
[[PAPER:2330|1]]
[[PAPER:2331|1]]
[[PAPER:2332|1]]
[[PAPER:2333|1]]
[[PAPER:2337|1]]
[[PAPER:2334|1]]
[[PAPER:2335|1]]
[[PAPER:2336|1]]
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
This species page was written by Kelly Ivors and Matt Greene (NC State University), with picture contributions from Kelly Ivors, Matt Greene, Larry Grand and D. Mike Benson (NC State University) and Daniel Huberli (Murdoch University).