
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
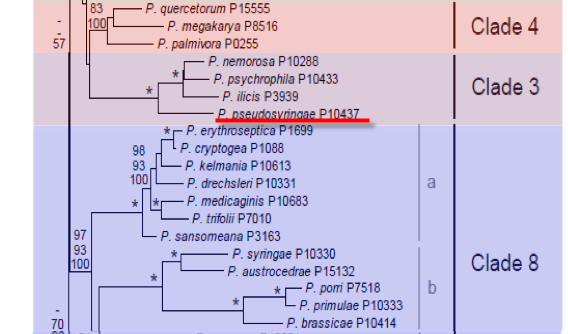
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora pseudosyringae T. Jung & Delatour 2003 (Oomycetes, Pythiales)
Notes: This species has previously been frequently misidentified as Phytophthora syringae (Jung et al. 2003).
Substrate: Soil, roots; also fruits when inoculated.
Disease Note: Root and collar rot. A moderately aggressive pathogen of various Quercus spp. (Jung et al. 2003); also causing a fruit rot of apple and leaf rot of Ilex aquifolium when inoculated.
Host: Quercus spp., Fagus sylvatica, Alnus glutinosa and other hardwood species.
Supporting Literature:
Jung, T., Nechwatal, J., Cooke, D.E.L., Hartmann, G.C., Blaschke, M., Oswald, W.F., Duncan, J.M., and Delatour, C. 2003. Phytophthora pseudosyringae sp. nov., a new species causing root and collar rot of deciduous tree species in Europe. Mycol. Res. 107: 772-789
Updated on Aug 07, 2006
Phytophthora pseudosyringae T. Jung & Delatour was initially recovered from soil in oak decline forests and necrotic fine feeder roots and stem necrosis of Fagus sylvatica and Alnus glutinosa. Phylogenetically it is a clade 3 species closely related to P. ilicis, P. nemorosa, and P. psychrophila.
1. Sporangia
Sporangia were produced rarely on solid agar and abundantly in water culture. They were borne terminally on unbranched sporangiophores or more often in irregular lax or regular dense sympodia. Sporangia were semi-papillate, very rarely bi-papillate, and usually limoniform, ellipsoid or ovoid. Obovoid, pyriform, obpyriform or distorted shapes were less common. Caducous sporangia with pedicel lengths of <5–20 µm were present in all isolates, but their proportion varied between isolates from 10 to >80%. Sporangia averaged 47.5 ± 6.3 x 29.5 ± 3.2 µm (overall range 20–98 x 15–48 µm) with a wide range of isolate means of 38–66 x 23–41 µm, and a length/ breadth ratio of 1.61 ± 0.13 (range of isolate means 1.43–1.96). The two extremes were represented by the alder isolate PSEU 17 (38±7 x 23±3.8 µm) and isolate PSEU 12 from oak (66 ± 9.9 x 34 ± 5 µm). Sporangia with unusual features such as markedly curved apices, a conspicuous basal plug or lateral attachment of the sporangiophore were common in some isolates and rare in others. Zoospores were discharged through an exit pore 4–10 µm wide (av. 7.5 ± 0.5 µm). They were limoniform to reniform whilst motile, becoming spherical on encystment. Direct germination of sporangia was common in older water cultures
2. Chlamydospores
No chlamydospores were observed on all agar media
3. Sex Organs
Oogonia were readily produced in single culture on V8A, CMA-b and CA by most isolates; however, during storage some isolates gradually lost the ability to produce oogonia. Oogonia were smooth walled, spherical or rarely slightly ovoid, and borne terminally. Oogonial diameters from 15 isolates ranged from 16.5–42.5 µm and averaged 29 ± 2.9 µm with isolate means ranging from 24–34 µm. Oospores diameters of 14–21-day-old oospores ranged from 15–36.5 µm (av. 26.5 ± 2.8 µm). Oospores were spherical, usually plerotic or almost so. Oospore walls averaged 1.35 ± 0.22 µm (0.5–2.5 µm). In some isolates older oospore walls turned golden yellow. High levels of oospore abortion were found in all isolates. Antheridia were hyaline, single, terminal, and spherical or club-shaped to irregular. Their dimensions ranged from 8.5 to 26 x 6.5 to 15 µm (av. 14.8 ± 9.5 µm). They were predominantly paragynous (>95%), usually inserted near the oogonial stalk, but some amphigynous ones were also present in most isolates.
4. Growth Temperatures
At 20° C colonies grew moderately slowly on V8A, CA, MEA, CMA-b with fastest growth on V8A and very slowly on PDA. Growth on V8A occurred from 5 to 25° C with an optimum at 20° C (radial growth rate 4.6 ± 0.8 mm/day). No growth occurred at 27.5 and 30° C. All isolates showed re-growth after 7 days at 27.5° C when returned to 20° C, whereas 30° C was lethal for all isolates.
5. Growth Characteristics in Culture
Colony growth patterns of 18 isolates showed some variation, but were generally stellate on V8A and CA, and stellate to rosaceous on MEA and CMA-b, appressed to limited aerial on CA and CMA-b and limited aerial (with fluffy patches) to fluffy on V8A and MEA. Cultures of isolates PSEU 25 and 28 were striate rather than stellate on CA, MEA and CMA-b, whereas the isolates PSEU 13, 15, 17 and 29 from alder produced more aerial mycelium on V8A, CA and MEA than the isolates from oak and beech. On PDA colonies of all isolates were appressed, and stellate to rosaceous patterns were produced. Primary hyphae on V8A were 4–9 µm wide (average 6.2 µm) and did not branch sympodially. Irregular to coralloid lateral hyphae were common in most isolates. 17 out of 23 isolates produced an orange-brown pigment on CHT agar. All 3 isolates from alder and 3 of the 17 isolates from oak failed to produce pigments.
Chains of inflated, spherical to deltoid hyphal swellings, sometimes with radiating hyphae, were abundantly produced in water culture by all isolates from oak and beech tested and averaged 17.5 ± 3 µm (7.5–32.5 µm). All three isolates from alder (PSEU 13, 17 and 29) formed only small numbers of single spherical swellings.
6. Distinguishing Characteristics
P. ilicis is the closest relative to P. pseudosyringae. Nevertheless, it is easily distinguished from P. pseudosyringae by the absence of paragynous antheridia, smaller oogonia, the production of sporangia on solid agar, the absence of catenulate hyphal swellings in water culture, different colony growth patterns on V8A, CA, MEA, CMA-b and PDA, lower maximum temperature for growth, and a different pigmentation on CHT agar. Furthermore, P. ilicis occurs exclusively on above-ground parts of Ilex aquifolium.
P. psychrophila is distinguished by having exclusively amphigynous antheridia, larger oogonia, sporangia with much more variable, often distorted shapes, that are also produced at the surface of and submerged in solid agar, by sympodial branching of hyphae, the absence of catenulate hyphal swellings in water culture, different colony growth patterns on V8A, CA, MEA, CMA-b and PDA, lower optimum and maximum temperatures for growth, slower growth rates, different pigmentation on CHT agar, and a difference in ITS (6 bp) and mtDNA sequences. Furthermore, P. psychrophila was less aggressive to fine roots of oaks, non-pathogenic to leaves of English holly and apple fruits, and was never recovered from necrotic bark of beech and alder.
P. quercina differs from P. pseudosyringae by the absence of amphigynous antheridia, a large proportion of elongated oogonia, thicker oospore walls, distinctly papillate persistent sporangia with much more variable shapes, sympodial branching of hyphae, the absence of catenulate hyphal swellings in water culture, different colony growth patterns on V8A, CA, MEA, CMA-b and PDA, higher optimum and maximum temperatures for growth, slower growth rates, different pigmentation on CHT agar, and different ITS and mtDNA sequences. Furthermore, P. quercina is markedly more aggressive to fine roots of pedunculate oak and almost non-pathogenic to fine roots of beech, leaves of English holly and apple fruits.
P. nemorosa causes lethal stem cankers on Lithocarpus densiflorus and Q. agrifolia and foliar lesions on L. densiflorus and Umbellularia californica in California and Oregon. According to ITS sequence analysis it is also closely related to P. ilicis, but differs from P. pseudosyringae by having larger oogonia and oospores with exclusively amphigynous antheridia, and by globose to broadly-ovoid sporangia with a markedly lower l/b ratio (1.32 vs 1.61). In addition, P. nemorosa forms blistered instead of deltoid catenulate hyphal swellings, has lower maximum and optimum temperatures for growth, and is non-pathogenic to English holly.
P. hibernalis is another semi-papillate low-temperature species that differs from P. pseudosyringae by the predominantly amphigynous antheridia, the production of smaller sporangia (average 40 x 19 µm) with a tapering base and longer pedicels (23–73 µm) when caducous, by lower maximum and optimum temperatures for growth, different colony growth patterns, and by putatively different host ranges.
P. syringae, despite being one of the earliest described Phytophthora species, has often been misidentified resulting in a series of invalid synonyms. In particular, P. hibernalis and P. syringae have often been confused. Also many isolates of P. pseudosyringae have erroneously been identified as P. syringae. The reasons for these misidentifications were similarities with morphological and physiological properties assigned to P. syringae which were confirmed by this study, i.e. predominantly paragynous antheridia, semi-papillate sporangia that are borne in sympodia, inflated catenulate hyphal swellings in water culture, low cardinal temperatures for growth and moderately slow growth rates. Nevertheless, P. syringae can easily be distinguished from P. pseudosyringae by having persistent sporangia, that are abundantly produced also on solid agar, with a markedly lower l/b ratio (1.43 ± 0.1 vs 1.61 ± 0.1) and wider exit pores (9.5¡0.4 vs 7.5¡0.4 µm), slightly different colony growth patterns on V8A, CA, MEA and CMA-b, lack of growth on PDA, lack of pigmentation on CHT agar, and clearly different ITS sequences (83% similarity).
P. citricola also shows some similarities to P. pseudosyringae, i.e. paragynous antheridia, semi-papillate sporangia and stellate growth patterns on V8A and CA. However, P. citricola is clearly separated by the absence of caducous sporangia and hyphal swellings in water culture, different colony growth patterns on CMA, MEA and PDA, higher optimum (27.5 C) and maximum (>30° C) temperatures for growth and much faster growth rates (8.1 and 9.5 cm/day at 20 and 25° C). Furthermore, P. citricola has quite different ITS sequences and is markedly more aggressive to Fagus sylvatica and Alnus glutinosa
Causes root rot of Quercus robur, Q. petraea and Acer platanoides, root rot and bark and stem lesions on F. sylvatica, foliar lesions on Umbellularia californica.
Jung, T., Nechwatal, J., Cooke, D. E. L., Hartmann, G., Blaschke, M., Oßwald, W. F., Duncan, J. M. and Delatour, C. 2003. Phytophthora pseudosyringae sp. nov., a new species causing root and collar rot of deciduous tree species in Europe. Mycol. Res. 107: 772–789
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS. This species page was adapted from Jung et al. (2003)
Isolate list