
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
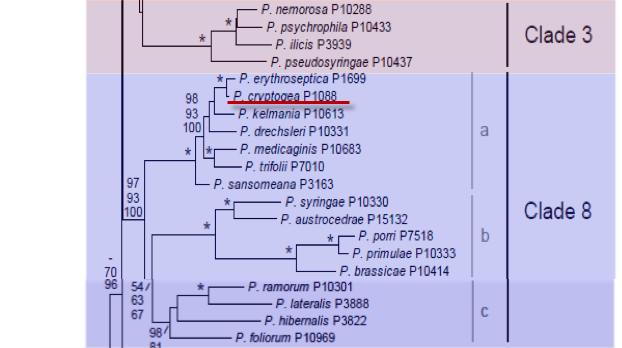
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora cryptogea Pethybr. & Laff. 1919 (Oomycetes, Pythiales)
Notes: Ho and Jong (1986) considered Phytophthora cryptogea to be synonymous with Phytophthora drechsleri, Cooke et al. (2000) found the two species to be distinct based on molecular analysis, but close to Phytophthora erythroseptica.
Distribution: Cosmopolitan.
Substrate: Roots, stems, corms, leaves.
Disease Note: Damping-off, foot rot, stem rot, leaf rot, wilt. See Stamps 1978.
Host: 141 genera in 49 families.
Supporting Literature:
Cooke, D.E.L., Drenth, A., Duncan, J.M., Wagels, G., and Brasier, C.M. 2000. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genet. Biol. 30: 17-32
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Ho, H.H., and Jong, S.C. 1986. A comparison between Phytophthora cryptogea and P. drechsleri. Mycotaxon 27: 289-319
Stamps, D.J. 1978. Phytophthora cryptogea. C.M.I. Descript. Pathog. Fungi Bact. 592: 1-2
Updated on Jun 06, 2006
P. cryptogea is morphologically very similar to P. drechsleri. Differentiation of P. cryptogea from P. drechsleri is discussed in Chapter 4 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). The major distinguishing feature that separates P. cryptogea from P. drechsleri is the growth of P. drechsleri isolates at 35oC (Tucker 1931). See Table 1 for a summary of classical differences between the two species. Bumbieris (1974) studied the morphology and physiology of both P. cryptogea and P. drechsleri and concluded that the species were identical; he suggested that P. drechsleri should be included within P. cryptogea, since this species was described earlier. Similar conclusions were reached by Hallsall (1976), who found isolates of both species to be serologically related; by Shepherd (1978) on the basis of mating behavior; by Matsumoto and Sato (1979), who noted that the two species had similar protein patterns; and by Ho and Jong (1986), who examined the morphology and physiology of a large number of species described as either P. cryptogea or P. drechsleri. Ho and Jong found that the sporangial shapes (tapered or rounded bases) and sizes of these two species varied considerably with much overlapping of characteristics, making it difficult categorically to separate P. cryptogea from P. drechsleri. An isolate from bean produced sporangia with both tapered and rounded bases (Flowers et al. 1973). In a later report, Ho and Jong (1991) decided that both species should be retained and differentiated by growth at 35oC for P. drechsleri and no growth for P. cryptogea isolates, a character used by Tucker (1931). Protein profiles obtained by polyacrylamide gel electrophoresis (see Chapter 4 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996) showed that a type culture of P. cryptogea and an authentic culture of P. drechsleri were clearly differentiated. Several other isolates with similar morphology had protein profiles intermediate between those of P. cryptogea and P. drechsleri, thus making it impossible to resolve the identities of these isolates with either of the above pecies (Cacciola et al. 1990). The protein profile was distinctly different from that of P. erythroseptica. An isolate from lentisk yielded a protein profile identical to that of the type culture of P. cryptogea and was considered to be P. cryptogea (Magnano di San Lio 1992). If 35oC is valid temperature for differentiation, the causal agents of certain diseases listed in Table 1 would have to be revised. For example, the causal agent of Phytophthora root rot of safflower grows at 35oC; however, the name of some of these isolates was changed from P. drechsleri to P. cryptogea by Klisiewicz (1977) on the basis of production of sporangia with rounded bases (Figure 23.1 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). P. cryptogea is assigned to taxonomic group VI of the genus (Waterhouse 1963; Newhook et al. 1978; Stamps et al.1990). The morphology of the asexual and sexual structures of this species is illustrated in Figure 1 and Figure 2. See Chapter 4, Tables 4.2 and 4.3 for tabular keys (data of Ho and Jong [1991]), Appendix 4.9 for a dichotomous key (Ho 1992) in Phytophthora Diseases. Recently, Mills et al. (1991) studied the isozyme and mtDNA RFLP patterns of 123 isolates designated either P. cryptogea or P. drechsleri that had been isolated from various host plants from the United States and overseas. Their findings indicated that groups within these two species varied greatly, and consequently they concluded that merging P. cryptogea with P. drechsleri was not warranted. Instead, their data indicate that there are at least seven distinct genetic groups within these two species. Some of these genetic groups differ more in mtDNA patterns than do some accepted species. A revised description of P. cryptogea and P. drechsleri recently presented by Ho and Jong (1991) broadened the description of P. cryptogea to include isolates that are intermediate between P. cryptogea and P. drechsleri and retained the criterion of growth of P. drechsleri at 35oC to separate it from P. cryptogea. In the report by Mills et al. (1991) on isozymic and mtDNA RFLP patterns, all isolates in group A grew at 35oC. However, another group, H, which differed genetically from group A, also grew at 35oC. Isolates from pigeon pea, named P. cajani or P. drechsleri f. sp. cajani, which are included in P. drechsleri because of growth at 35oC, also were genetically different from all other groups (Mills et al. 1991). So these two studies agree in some ways but not in others that growth at 35oC is a critical criterion. See Table 2 for a summary of characteristics recorded for P. cryptogea and P. drechsleri, and Table 3 for dimensions of spores recorded in different references. The amended description by Ho and Jong (1991) is as follows (reproduced by permission of Mycotaxon Ltd., Ithaca, New York, courtesy of Ho and Jong 1991): Mycelium with no distinct or slightly floral growth pattern on clarified V-8 juice agar medium, slightly fluffy to fairly fluffy; aerial mycelium often profuse on unfiltered yellow corn meal agar medium; main hyphae uniform to uneven 5µm wide, commonly forming in water; conspicuous network of small hyphal swellings (under 25µm), spherical to irregular. Sporangia rarely produced on agar but abundantly in water, nonpapillate, nondeciduous, internally proliferating, terminal on unbranched or often close sympodially branched sporangiophores (1-3 µm wide); first-formed sporangia regularly obpyriform to ovoid, mostly with rounded base but later-formed sporangia tend to be more elongated, asymmetrical or variable in shape; sporangia av. 52 x 30 µm (35-63 x 24-35 µm), L/B ratio 1.7 (1.4-2.3), apex flattening on mounting; sporangia collapse after zoospore release which often requires chilling; exit pore over 8 µm wide, pore/breadth ratio over 0.3. Heterothallic though sex organs sometimes formed sparsely in aged culture; oogonia spherical to subspherical, smooth, av. 31 µm (28-37 µm) diam., wall becoming yellowish to brown; oospores plerotic, spherical, av. 27 µm (24-32 µm) diam., wall 2-4µm thick; antheridia amphigynous, short cylindrical, 14 µm (12-17 µm) x 14 µm (13-17 µm), unicellular. No chlamydospores formed in culture. Cardinal temperatures for growth: 0-5oC, 20-25oC, 30-35oC; growth rate on clarified V8 agar medium 5-10 mm per day at 20oC. Until more research is conducted to identify the numerous genetically different biotypes, the description as amended by Ho and Jong (1991) should be used to characterize P. cryptogea. See Chapter 23, P. drechsleri, in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996), for a more complete discussion of the P. cryptogea-P. drechsleri complex. It would appear doubtful that isolates that grow at 35oC but otherwise have morphological features similar to those of P. drechsleri or P. cryptogea should be classified as P. cryptogea, e.g., P. cryptogea on safflower (Carthamus tinctorius) (Table 3).
1. Sporangia
Sporangia are oval to obpyriform with rounded bases, nonpapillate, and persistent on the stalk. Sporangia that develop later tend to be more elongated or variable in shape (Ho and Jong 1991). Dimensions of sporangia range from 20 to 93 µm in length and 15 to 51 µm in breadth, with average measurements of 37 to 40 x 23 µm (Stamps 1978b). Measurements of sporangia from various host plants are given in Table 20.2. As stated in Chapter 3, nutritional status has a great effect on the size of sporangia. In Table 4, the ranges of sizes given in various reports show that sporangia grown initially on potato-dextrose agar are smaller than those grown on pea broth. Thus, comparison of sporangial sizes is of doubtful value for taxonomic work unless the cultural conditions (e.g., nutrients in the substrate), the type of washing solution used on mycelium (e.g., water, mineral salts, or soil extract), and incubation under light are the same. See Chapter 3 in Erwin and Ribeiro (1996, Phytophthora Diseases Worldwide) for a discussion of sporangial formation. The length-breadth ratio is 1.45:1 to 1.57:1. Ho and Jong (1991) report ratios of 1.4 to 2.3 with an average of 1.7. Sporangia rarely form in solid culture media but form in abundance in a suitable aqueous substrate. See Chapter 3 for methods of inducing sporangia. Sporangiophores are 2 to 3.5 µm in width and proliferate through the empty sporangium (Figure 2d) or develop sympodially from below the sporangium (Figure 1).
2. Chlamydospores
Chlamydospores are not usually known to occur; however, chlamydospores formed on pine roots in soil (Bumbieris 1971).
3. Sex Organs
P. cryptogea is heterothallic and requires pairing with an opposite mating type to form oogonia and oospores. Antheridia are amphigynous and spherical and average 10 µm in diameter. Antheridia may sometimes be oval or cylindrical, depending on the species used for the mating. Oogonia rarely form in single culture. Oogonia have tapered bases and measure 28 to 40 µm in diameter with an average of 30 µm. They are smooth walled and become yellow/brown with age. Oospores are plerotic or nearly so, measure 20 to 32 µm in diameter with an average of 26 µm, and are thick-walled (3.5 µm) (Figure 1).
4. Growth Temperatures
The minimum temperature for growth is <1oC; optimum temperature for growth is 22 to 25oC; and the maximum temperature for growth is 31 to 33oC. Temperature relations of isolates from various hosts (Table 2) show that there are variations from these criteria, particularly at minimal temperatures.
5. Cultural Characteristics
Cultures are uniform, with some isolates displaying a slight to distinct floral pattern. Mycelium in V8 juice agar medium is fairly fluffy.
6. Distinguishing Characteristics
The hyphae are very uneven in width. In liquid culture, characteristic angular hyphal swellings form. Hyphal swellings are rarely observed in solid culture media. Hyphal swellings form in chains or clusters and average 11 µm in diameter with a maximum of 20 µm (Figure 1 and Figure 2c). No chlamydospores are observed in culture media. Although they are not common, chlamydospores have been reported for some P. drechsleri isolates (Cother and Griffin 1973b, 1974). Kröber(1981a, b) described the forma specialis begoniae of P. cryptogea on the basis of host specificity to begonia. The sporangia of P. cryptogea f. sp. begoniae averaged 46.7 x 30.5 µm; oogonia averaged 30.8 µm; oospores averaged 28.3 µm in diameter; and hyphal swellings averaged 20.8 µm. This forma specialis was A2 mating type. Isolates from begonia formed a group D when assayed for isozyme patterns (Mills et al. 1991). Some isolates differ in pathogenicity to different hosts. For example, the cultivar Transvaal daisy isolates are pathogenic to Mattiola sp. and Callistephus chinensis only, whereas isolates from Callistephus are pathogenic to both Transvaal daisy and Mattiola sp. as well as cucumber, Cheiranthus, eggplant, tomato, Capsicum sp., pumpkin, and watermelon (Tompkins and Tucker 1937). Isolates from Gloxinia sp. (Middleton et al. 1944) and chicory (Forlot et al. 1966) are not host specific.
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Isolate list