
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
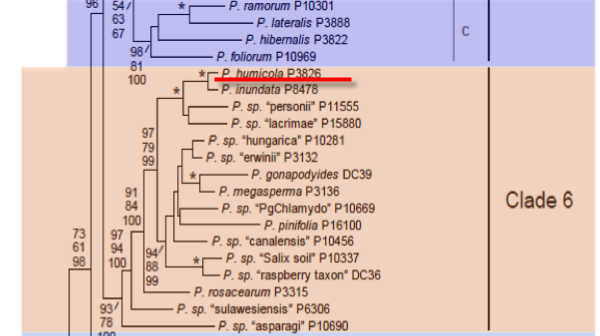
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora humicola W.H. Ko & Ann 1985 (Oomycetes, Pythiales)
Distribution: Asia (Taiwan, type).
Substrate: Soil. Able to infect fruits, roots, leaves.
Disease Note: Pathogenic to apple and cucumber, but not orange or eggplant, after wound inoculation.
Host: Natural host unknown, but isolated from a Citrus sp. orchard (Rutaceae). Inoculated hosts include Malus domestica (apple, Rosaceae) and Cucumis sativus (cucumber, Cucurbitaceae).
Supporting Literature:
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Kroon, L.P.N.M., Bakker, F.T., van den Bosch, G.B.M., Bonants, P.J.M., and Flier, W.G. 2004. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNS sequences. Fungal Genet. Biol. 41: 766-782
Updated on Jun 13, 2006
P. humicola is classified in group V (Stamps et al. 1990). See Tables 4.2 and 4.3 for tabular keys and Appendix 4.9 for a dichotomous key (Ho 1992) in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996). Morphology is shown in Figure 1. Data presented below are from Ko and Ann (1985).
1. Sporangia
Sporangia are formed only when pieces of newly colonized V8 juice agar are incubated in nonsterile soil extract under diffuse light. Incubation in water or other conventional salt solutions does not induce sporangial formation. Nonpapillate, ovoid sporangia, measuring 38 to 60 µm x 50 to 72 µm (average 44 - 58 µm), are noncaducous. Zoospores are released after the temperature of the sporangia in soil extract is reduced to 8°C for 30 min. Sporangia proliferate internally but rarely externally. They occasionally are produced on the tips of hyphae radiating from germinating hyphal swellings.
2. Chlamydospores
Chlamydospores have not been observed although large, spherical hyphal swellings measuring 22 to 56 µm, average 40 µm, are produced in sterile distilled water. Radiating hyphae emanate from the hyphal swellings. Sometimes the dominant hypha produces another hyphal swelling and forms a chain of hyphal swellings.
3. Sex Organs
P. humicola is homothallic. About 47,000 oospores per square centimeter were produced on the surface of a petri plate of 20% V8 juice agar incubated at 25 to 28°C in the dark. Oospores were not produced at 23°C or below or above 30°C. Continuous light prevented oospore formation. Oogonia are spherical and smooth walled, measure 25 to 52 µm in diameter, average 39 µm, and are borne on long stalks. Antheridia are mostly paragynous and ovate to elliptical or irregular in shape and are borne on stalks 8 to 89 µm, average 31 µm, in length. Amphigynous antheridia occur but are rare. Paragynous antheridia measure 10 to 16 µm - 11 to 26 µm, average 14 - 18 µm, and amphigynous antheridia measure 15 to 19 - 15 to 21 µm, average 16 - 17 µm.
4. Growth Temperatures
The minimum temperature for growth is 9°C, optimum 28 to 30°C, and maximum 32°C.
5. Distinguishing Characteristics
Ko and Ann (1985) state that P. humicola resembles P. megasperma because of the production of nonpapillate, noncaducous sporangia that proliferate internally and the relatively large oogonia and oospores with predominantly paragynous antheridia (see Chapter 46). They state that the production of large hyphal swellings and the long stalks on oogonia and antheridia distinguish P. humicola from P. megasperma and that the protein pattern (by disk gel electrophoresis) of P. humicola is distinct from that of P. megasperma f. sp. glycinea (now known as P. sojae [Hansen and Maxwell 1991]). However, P. sojae is now known to not be a member of the P. megasperma species complex [see Chapter 46 in Phytophthora Diseases Worldwide (Erwin and Ribeiro 1996)], therefore these data do not prove a difference from P. megasperma. However, use of molecular restriction fragment length polymorphisms (RFLP) on mitochondrial DNA (mtDNA) provide data that have shown a marked variability in the genetic makeup of various biotypes within P. megasperma (Förster and Coffey 1993). A comparison of the mtDNA RFLP pattern of P. humicola with the different types of P. megasperma that have been described would provide more convincing evidence that P. humicola is distinct.
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Isolate list