
Phytophthora has been rebuilt to fix security-related problems and to restore GIS tools. These tools allow users to visualize the geospatial, temporal, and environmental contexts of Phytophthora discoveries. The next phase is to update species information and add data derived from large-scale surveys. If you have suggestions and requests to make the database better, please contact Seogchan Kang (sxk55@psu.edu).
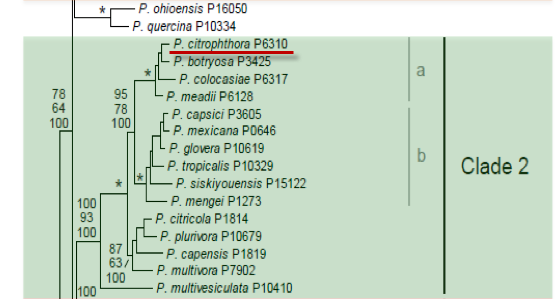
Genus wide phylogeny for Phytophthora using four mitochondrial loci (cox2, nad9, rps10 and secY; 2,373 nucleotides). Maximum likelihood branch lengths shown. Numbers on nodes represent bootstrap support values for maximum likelihood (top), maximum parsimony (middle) and Bayesian posterior probabilities as percentages (bottom). Nodes receiving significant support (>95%) in all analysis are marked with an asterisk (*). Scale bar indicates number of substitutions per site.(Martin, Blair and Coffey, unpublished).

[ Click the tree to enlarge it. ]
Phytophthora citrophthora (R.E. Sm. & E.H. Sm.) Leonian 1906 (Oomycetes, Pythiales) Pythiacystis citrophthora R.E. Sm. & E.H. Sm. 1906
Pythiacystis citrophthora R.E. Sm. & E.H. Sm. 1906
Distribution: Cosmopolitan.
Substrate: Roots, stems, bark of trunk, twigs, leaves, fruits, pods.
Disease Note: Serious gummosis of citrus trees; in historical outbreaks nearly all citrus trees were destroyed (Erwin & Ribeiro 1996). Also a root rot, stem necrosis, canker, fruit rot, twig blight, seedling blight; see Erwin & Ribeiro for disease symptoms by host.
Host: 88 genera in 51 families.
Supporting Literature:
Erwin, D.C., and Ribeiro, O.K. 1996. Phytophthora Diseases Worldwide. APS Press, St. Paul, Minnesota, 562 pages.
Ho, H.H., and Chang, H.S. 1992. A re-evaluation of Phytophthora species described by K. Sawada in Taiwan. Mycotaxon 43: 297-316
Kroon, L.P.N.M., Bakker, F.T., van den Bosch, G.B.M., Bonants, P.J.M., and Flier, W.G. 2004. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNS sequences. Fungal Genet. Biol. 41: 766-782
Waterhouse, G.M., and Waterston, J.M. 1964. Phytophthora citrophthora. C.M.I. Descript. Pathog. Fungi Bact. 33: 1-2
Updated on Jun 06, 2006
P. citrophthora is classified in group II (Stamps et al. 1990). Morphology is shown in Figure 1. Tabular keys are presented in Tables 4.2 and 4.3 in Chapter 4 and a dichotomous key in Appendix 4.9 (Ho 1992). A summary on the morphology of P. citrophthora and diseases caused is reported by Waterhouse and Waterston (1964c). Mchau and Coffey (1994b) reported results of a study of 77 isolates from a world collection and redescribed the species. The following redescription of P. citrophthora is reproduced by permission from Mchau and Coffey (1994b):
Sporangia produced in sterilized 1% soil extract, also on solid agar by a few isolates. Extremely variable in shape; ellipsoid, broadly ovoid, globose, limoniform, or extremely distorted with prominent papillae and some semipapillate in same isolates: 27  3.4-65.3 13.9 x 18.9 1.8-40.4 5.6 µm; length:breadth ratio 1.3:1 0.2 to 1.8:1 0.1; sporangia often with 2 or more papillae. In soil extract, sporangia borne singly or in very loose sympodia of 2 sporangia, often laterally attached to sporangiophore; sporangiophore often with globose swellings at branching points. Non-caducous.
3.4-65.3 13.9 x 18.9 1.8-40.4 5.6 µm; length:breadth ratio 1.3:1 0.2 to 1.8:1 0.1; sporangia often with 2 or more papillae. In soil extract, sporangia borne singly or in very loose sympodia of 2 sporangia, often laterally attached to sporangiophore; sporangiophore often with globose swellings at branching points. Non-caducous.
Hyphae smooth or coarse, 3 µm diameter. Colonies dense cottony like, or rosette, or stellate, or patternless on V8C agar at 24o. Radial growth rate on V8C agar at 24o, 7.6 2.4 mm d-1 [per day]. Hyphal swellings present in some isolates, spherical to ovoid 1-6 1.2 µm diameter, some irregularly shaped, forming within 3-4 days at 15o in sterilized 1% soil extract water. Stasis at 5o, optimum growth at 18-7o, lethal at 35o on V8C agar in the dark.
Chlamydospores present in some isolates, terminal or intercalary, spherical, 10-3 5.2 µm diameter some with irregular shapes, hyaline, forming within 7 days, walls 0.5-.5 µm thick when produced in V8C agar in darkness at 25o.
Sterile, though can sometimes induce oospore formation in other heterothallic species, e.g., A1 mating types of P. capsici. Isozyme phenotype. Monomorphic in MDH2, PGD1, PGD2, IDH2, LDH2, ME, FDP2, PGM, AK, and SOD. Polymorphic up to 25% in ACO, GPI, MDH1, IDH1, G6PDH, HEX2, and PEP. Paratypes: P318, P451, P1200, P1320, P1323, P1837, P3521, P3527, P3771, P3934, P6295, P6310, P7481.
1. Sporangia
Sporangia are mostly papillate, persistent, and not caducous. Although sporangia are described by Stamps et al. (1991) to be caducous in some isolates but not in others, Mchau and Coffey (1994b) reported that all of 77 isolates produced noncaducous sporangia. The shapes are variable, ranging from spherical, ovoid, obpyriform, obturbinate, to ellipsoidal; dimensions are 23 to 90 x 18 to 60 µm, average 45 x 30 µm. The length-breadth ratio is 1.2:1 to 2.0:1, average 1:4. Mchau and Coffey (1994b) report length-breadth ratios of 1.3:1 to 1.8:1 with a length range of 27.4-4.3 µm and a breadth of 18.9-0.4 µm. See Table 1 for a range of dimensions in other reports. Sporangia are occasionally laterally attached and some are intercalary. Sporangiophores are irregularly branched, some singly and some in a loose sympodium with a swelling at the point of branching.
2. Chlamydospores
Chlamydospores do not ordinarily occur on citrus isolates; however, chlamydospores formed in citrus roots from a California orchard during the warm summer months (Sjoerdsma et al. 1988). The diameter was 25 to 35 µm, average 28 µm. Mchau and Coffey (1994b) report chlamydospores only on isolates from cacao in Brazil. Chlamydospores were 10-3 µm in diameter and hyaline and were produced both singly and in clusters.
3. Sex Organs
In general, it has been assumed that the oospore stage is not a part of the life history of P. citrophthora. Sex organs do not occur in nature on citrus. Sex organs are not included in current keys (Stamps et al. 1990) and were not noted in the original description. We have illustrated oospore formation here because, experimentally, oospores have been shown to form when certain P. citrophthora isolates are paired on carrot agar. Four isolates from cocoa black pod in Brazil formed oospores when paired with an A2 isolate from citrus (P318 from Australia), and one cocoa isolate formed oospores when paired with P717 (A1), a P. citrophthora isolate (host unknown) from New Zealand. None of the cocoa isolates produced oospores when crossed among themselves, however, although some were A1 and another was A2 mating type. Kellam and Zentmyer (1986) concluded from these data that the compatibility or mating system was probably more complex than the usual A1/A2 system in Phytophthora.
Mchau and Coffey (1994b) report that less than 30% of the 77 P. citrophthora isolates studied produced oospores when paired with an A2 mating type of P. capsici. By use of the polycarbonate membrane method of Ko (1978; see Chapter 3), P. citrophthora isolates acted only as inducers of oospores in the P. capsici partners. Although oospores from pairing A1 and A2 isolates of P. citrophthora were reported by Savage et al. (1968), no evidence of oospores from pairing of P. citrophthora isolates was found. When isolates are paired with A1 or A2 mating types of other species, antheridia are amphigynous and oogonia are 26 to 36 µm in diameter and contain oospores 21 to 28 µm in diameter.
4. Growth Temperatures
The minimal temperature for growth is 5oC, the optimum 24 to 28oC, and the maximum 32 to 33oC.
5. Distinguishing characteristics
An isozyme pattern study of 32 isolates of P. citrophthora from various hosts, including citrus, cocoa, Actinidia deliciosa, rubber, walnut, strawberry, pistachio, Ficus elastica, Prunus domestica, Amygdalus communis, Ribes sanguineum, and Coptus japonica, indicated that P. citrophthora was genetically related to P. capsici, but within the P. citrophthora isolates studied, two subgroups, CITR1 and CITR2, could be defined. Isolates from citrus grouped into CITR1, and those from cocoa grouped into CITR2 (Oudemans and Coffey 1991a). In a subsequent report, a group CIT5 (P. citricola from avocado) showed a close relationship with CITR groups (Oudemans and Coffey 1993). By use of polymerase chain reaction amplification of rDNA, Lee and Taylor (1992) showed a close relationship between cocoa isolates of P. capsici (formerly P. palmivora MF4) and P. citrophthora.
Mchau and Coffey (1994b) found that three subgroups of P. citrophthora could be differentiated isozymically. Subgroup CITR1 contained isolates from a broad range of hosts, including citrus from various geographical areas. Subgroup CITR2 was composed exclusively of isolates from cacao grown in Brazil. Subgroup CITR3 included isolates from cacao grown in Indonesia.
Nomenclature information was provided by the the Systematic Botany and Mycology Laboratory in USDA-ARS.
Isolate list